 Webbed Reprint Collection
Webbed Reprint Collection
William H. Calvin
University of Washington
Box 351800
Seattle WA 98195-1800 USA
Email || Home
Page || publication list
|
William H.
Calvin
"The unitary hypothesis: A common neural circuitry for novel manipulations,
language, plan-ahead, and throwing?"
as it appeared in
Tools, Language, and Cognition in
Human Evolution, edited by Kathleen R. Gibson and Tim Ingold.
Cambridge University Press, pp. 230-250 (1993).
copyright ©1993 by William
H. Calvin and Cambridge University Press.
|
Abstract
Plan-ahead becomes necessary for those movements which
are over-and-done in less time than it takes for the feedback loop to
operate. Natural selection for one of the ballistic
movements (hammering, clubbing, and throwing) could evolve a
plan-ahead serial buffer for hand-arm commands that would benefit
the other ballistic movements as well. This same circuitry may also
sequence other muscles (children learning handwriting often screw
up their faces and tongues) and so novel oral-facial
sequences may also benefit (as might kicking and dancing). An
elaborated version of the sequencer may constitute a Darwin
Machine that spins scenarios, evolves sentences, and facilitates
insight by offline simulation. An example is given of an
evolutionary scenario from an apelike ancestor, demonstrating the
transition behaviors and growth curve considerations that any such
theory needs to have; this particular scenario (involving throwing
improvements) also suggests an explanation for the puzzling design
of the Acheulean "handaxe."
In considering transitions of organs, it is
so important to bear in mind the probability
of conversion from one function to
another....
Charles Darwin, On the Origin of Species, 1859
That evolution, over all-but-infinite time, could
change one physical organ into another, a leg into a
wing, a swim bladder into a lung, even a nerve net
into a brain with billions of neurons, seems
remarkable, indeed, but natural enough. That
evolution, over a period of a few million years,
should have turned physical matter into what has
seemed to many, in the most literal sense of the term,
to be some kind of metaphysical entity is altogether
another matter.
Derek Bickerton, Language and Species, 1990
Introduction
It is traditional to talk about language origins in terms of
adaptations for verbal communications, to talk about toolmaking in
terms of natural selection shaping up the hand and the motor
cortex, to talk about the evolution of intelligence in terms of how
useful versatile problem-solving and look-ahead would have been to
hominids. But might natural selection for one of these
improvements serve to haul along the others as well? Indeed,
might some fourth function be the seed from which the others
grew? What are the chances of coevolving talk, technique, and
thought?
Getting "something for nothing" is, I know, profoundly anti-Calvinist.
And the Puritan ethic seems to require us to look for a
function's antecedents in their usefulness to that very function. But,
as the epigram shows, new uses for old things is not anti-Darwinian.
Conversions of function can be profound, providing an
enormous boost toward a new ability. Capabilities occasionally
arrive unheralded by gradual predecessors. In the familiar case of
bird flight origins, one observes that it takes a lot of wing feathers
to fly even a little. Presumably natural selection for thermal
insulation shaped forelimb feathers up to the threshold for flight.
Natural selection for a better airfoil shaped feathers thereafter, but
the switch-over from the insulation function to the flying function
was presumably a surprise, leaving the protobirds to explore their
newfound gliding abilities rather as we might try to figure out a
holiday gift that arrived without an instruction manual.
Inventions are the novelties in evolution, though you'd think
that shaping-up streamlining was what it was all about, when
reading most of the literature. But adaptations are only
improvements on an existing function; what we're talking about
here is the invention itself before streamlining, and that's often a
matter of a Darwinian conversion of function. Nature does take
leaps, and such conversions of function are even faster than those
anatomical leaps envisaged by proponents of punctuated equilibria
and hopeful monsters.
Why might we expect one of these hominid improvements
(tools, language, intelligence -- and, I might add, accurate throwing
and making music) to serve as a preadaptation for the others? First
the general reason, then some specific ones.
In general, the brain is better at "new uses for old things"
than any other organ of the body. Sometimes two digestive
enzymes, each evolved separately for a different food, can act in
combination to digest a third foodstuff. But a brain can easily
combine sensory schemas and movement programs in new ways,
since it tends to use a common currency that allows disparate things
to be compared. From whatever source, an excitatory or inhibitory
input is first converted into positive or negative millivolts; nerve
cells then add and subtract in this substitute value system. For one
input to substitute for another, it is only necessary that they produce
similar voltage changes in the relevant nerve cells. One can add
apples and oranges to get so many pieces of fruit.
More specifically, the aforementioned functions are all
examples of stringing things together, that specialty of left
brain:
- For toolmaking and tool use, one usually needs to make a
novel sequence of movements.
- To go beyond the categories of ape language (chimpanzees
have about 36 standard vocalizations, with 36
associated standard meanings) and say something
novel, our ancestors seem to have hit upon the trick
of assigning meaning to a sequence of
individually-meaningless vocalizations.
- To create a novel plan of action, one needs to spin a
scenario connecting the past with the future.
And then contemplate it before acting (since most
novel courses of action are either nonsensical or
dangerous), perhaps going through a number of
rounds of variation and selection to improve its
prospects.
- This "get set" phase is particularly important for versatile
throwing; one has to produce trains of nerve
impulses going to dozens of muscles, timed like the
finale of a fireworks display sequence.
- And music is a time series much like language; instead of
phonemes with various frequency components, we
have chords. Instead of stringing phonemes together
to make words and sentences, we string chords
together to make phrases and melodies, build an
emotionally-satisfying structure in time and sound.
But why might all these functions overlap in the brain? Why might
some neural machinery be used interchangeably for one, then the
other?
"Borrowing" Neural Machinery
Thanks to our map-making tendencies, we usually assume
that the neural machinery for moving the hand should have little to
do with that for moving the mouth: at least on the motor strip,
they're in "different compartments" (though, I might note,
neighboring ones). But it is premotor and prefrontal cortex that has
the reputation for stringing things together, getting them ready to
finally execute, not motor cortex. It isn't even clear that such
sequences feed exclusively through motor strip, since premotor
cortex has as many direct connections to spinal cord as does motor
strip itself.
Furthermore, both inputs and outputs tend to be broadly
wired in cerebral cortex: the functional maps may suggest
segregation, but the anatomy is one big smear (another example of
Alfred Korzybski's "The map is not the territory"). For example, in
the somatosensory path from a fingertip to the cortex's sensory
strip, a given thalamocortical neuron responding only to that
small patch of skin will nonetheless send axon branches to nearly
all of the cortical map for the hand, not just the map of that
particular finger (presumably the synaptic strengths are stronger
there than elsewhere). And a cell near the thumb-face boundary
line of sensory strip seems to receive inputs from both; some weeks
the face connections are stronger, other weeks the hand connections
dominate that cell's functioning (references in Calvin, 1989).
A cell in motor cortex, seemingly specializing in a hand
movement, will nonetheless send axon branches to numerous other
segments of the spinal cord; perhaps overcoming the clumsiness of
youth involves tuning up the synaptic strengths to emphasize one
destination over the others. So to suggest that the parts of cerebral
cortex which optimize hand movements might also aid mouth
movements is not such a radical proposal. Indeed, Charles Darwin
made such neurological observations in his 1872 book, The
Expression of the Emotions in Man and Animals:
[Persons] cutting anything with a pair of
scissors may be seen to move their jaws
simultaneously with the blades of the scissors.
Children learning to write often twist about
their tongues as their fingers move, in a
ridiculous fashion.
It looks as if that broad smear of wiring may sometimes cause
simultaneous activation of rather different muscle groups. The
tendency of right-handed gestures to accompany left-brain-generated
speech (Poizner et al, 1987) is only the most commonly noticed
manifestation of this higher-order control of sequence.
Cortical Specializations for Sequence per
se
A hundred years after Darwin, the neuropsychologists also
began to emphasize sequencing. A. R. Luria noted (a modern
review is Stuss & Benson, 1986; pp. 225-226) how patients with
prefrontal damage had difficulty in planning a sequence of
movements. For example, a patient is in bed with his arms under
the covers. He is asked to raise his arm. He doesn't seem able to
do so. But if you ask him to remove his arm from under the
covers, he can do that. If you then ask him to raise it up and down
in the air, he does it all correctly and smoothly. There's no
paralysis (which would suggest damage to motor strip or below),
and no difficulty with executing a fluent sequence (which would
suggest premotor cortex damage) -- just a difficulty in
planning the sequence, getting stuck on the condition of
working around the obstacle of the confining bedcovers.
Further posteriorly, there was demonstrated an overlap
between language disorders and sequential hand movements. In
aphasic patients without any paralysis, Doreen Kimura (1979) and
her coworkers showed that these stroke patients had trouble
stringing together a sequence of hand movements, even though they
could carry out each individual motion with ease. To use an
everyday example (rather than the novel manual-brachial sequences
they tested), the patient might be able to turn a key in a lock, turn
the doorknob, and push open the door -- but not all in one facile
sequence. It was as if the stroke had damaged some sequencing
machinery that was often used for language but also for planning
novel hand-arm sequences.
And then the neurosurgeon George Ojemann began repeating
Wilder Penfield's classic brain stimulation studies in awake epileptic
patients, using much more sophisticated tests of language and
related sensory- and movement-sequencing functions (Ojemann,
1983; Ojemann & Creutzfeldt, 1987; Calvin & Ojemann, 1980).
"Language cortex" includes such left frontal lobe regions as
premotor and prefrontal, and extends posteriorly to include the
parietal and temporal lobe regions surrounding the Sylvian fissure.
While highly variable from patient to patient, the peri-Sylvian
"core" of the cortex from which function could be disrupted was
always related to sequencing. Tests of nonlanguage auditory
sequences were disrupted from the same regions as tests of
nonlanguage oral-facial movement sequences, suggesting that such
cortex subserves both receptive and expressive sequencing. This
"sequencing core" is typically surrounded by a peripheral region
where stimulation disrupts short-term verbal memory or some other
aspect of language.
So it would appear that the brain has some regions which
are particularly specialized for generating and analyzing sequences,
and that they may be used by multiple sensory and motor systems.
Why a Versatile Sequencer?
The left brain has some specializations for sequencing that
go back at least as far as the monkeys -- we don't know how it
evolved, but the left hemisphere seems to be better at deciphering a
novel string of sounds than is the right hemisphere (reviewed by
Falk, 1987). But there isn't much evidence that animal calls are
sequenced for an additional meaning, except for the escalating
loudness of monkey cries possibly indicating predator proximity.
On the motor side of things, it is easier to see why some
neural sequencing machinery might be needed. Normally, it is not
essential to make a detailed movement plan before starting to move.
That is because goal-and-feedback works pretty well, even for most
novel movements, as the brain gets "progress reports" along the
way. But there are some movements which are too rapid for
feedback to guide them: they are over-and-done-with by the time
that the first feedback arrives. Compared to most electronic and
hydraulic systems, neural feedback is quite slow: human reaction
times require hundreds of milliseconds. One of the fastest loops is
from arm sensors to spinal cord and back out to arm muscles: it
takes 110 milliseconds for feedback corrections to be made to an
arm movement (Cordo, 1987).
But dart throwing doesn't take much longer. So that
feedback from joints and muscles is wasted -- you might use it to
help you plan for the next time, but your arm is an unguided
missile shortly after the throw has begun. You must plan perfectly
as you "get set" to throw, create a chain of muscle commands, all
ready to be executed in exactly the right order. The same is true
for hammering, which both baboons and chimpanzees use
effectively to crack open shells. You need something like a serial
buffer memory in which to load up all the muscle commands in the
right order and with the right timing relative to one another -- and
then you pump them out blindly, without waiting for any feedback.
Recall player pianos, where you punch an instruction roll
that remembers which keys should be hit, and when. It may not
look like a sheet of music (it is 88 channels high, one for each key)
but it functions like one. The number of muscles that you need to
control for throwing or hammering is similar, once you start
counting up all those finger flexors and shoulder muscles.
To reprogram a player piano roll is a nuisance -- to make
corrections requires tape and a punch at a minimum -- and to slow
down an arpeggio in the middle of a long piece makes one wish for
a recording medium that could be locally stretched a little. Since
the brain is always doing little variations on a theme in improving
its hammering or throwing repertoire, it has probably found a less
awkward way of "recording motor tapes." Especially for throwing,
where the distance to the target and the desired speed tend to vary
widely with different targets -- and as a hunter, one is most often in
situations never encountered before.
More than One Planning Track for
Sequences?
As you get set to throw (at least in a situation less
standardized than a basketball free-throw), the brain has to discard
a lot of possibilities, narrow down the search to a few good
candidates, then select one to be let loose. So at least two serial
buffers are required, one to hold the current candidate train while it
is graded by memories of past successes and failures, and another
buffer to hold onto the best-so-far.
But the more planning tracks, the sooner a better candidate
sequence might emerge: think of dozens or even hundreds of
planning tracks in simultaneous operation. It would be like a
railroad marshalling yard, many candidate trains vying for a chance
to be let loose on the main track. You might keep the muscles
inhibited (much as in dreaming sleep) until you have found a good-enough choice, then you could "play that tape", acting out that
movement scenario (I suspect, however, that some reproduction
steps intervene). Until then, you presumably shuffle the trains-in-formation after each round, occasionally substituting something
linked in memory.
One obvious way for this to operate would be for the
higher-rated trains to "reproduce themselves" (with, of course,
copying errors on occasion) in the tracks of the trains rated poorly
by the memorized environment. In this manner, the "best
sequence" would also be the most common sequence (and attaining
a certain percentage of the total tracks might represent the threshold
for being the sequence being "let loose" on the main track of
action).
In Fig. x.1, I show words being sequenced, to use a serial-order example that is more easily communicated than muscle
command sequences. I show only four of the many planning
tracks, each randomly assembling a train of words, grading it
against memories of how similar sequences have performed in the
past, keeping the best one or two sequences, and trying again.
Such rounds of shaping up, as Dawkins (1986; ch. 3) showed for
shaping up random letter sequences to a Shakespearian model, can
quickly converge on a good-enough sequence.
The Brain as a Darwin Machine
This should be beginning to sound familiar. What other
process has lots of serially-coded individuals, the sequence judged
by the environment, and some individuals correspondingly more
successful at reproducing themselves (albeit with a little shuffling
of the code)? Actually, there are now two good examples: 1)
DNA strings called genes being shaped up by an
environment of prey-predators-pathogens, and 2) amino acid strings
called antibodies, shaped up during the immune response
by the differential reproduction associated with finding a foreign
protein to destroy. Shuffling the code a little seems to be far more
important than inducing errors, e.g., the permutations of
crossing-over during meiosis is the common source of new variants,
not mutations per se.
As I have recently argued (Calvin, 1987, 1989), many neural
sequencers to select among gives a very interesting property: brain
processes can, in effect, simulate the process used by Darwinian
evolution. They can shape up new serially-coded throws/thoughts/-
plans/sentences rather than new species or antibodies, using
innocuous remembered environments rather than real-time noxious
ones, operating in seconds rather than the days of
the immune response or the millennia of species evolution.
With this "Darwin Machine," the brain selects sequences of
schemas. Some possible examples:
- Any of the manual-brachial ballistic movements
(throwing, hammering, clubbing) might
benefit in situations where a standardized movement
could not be used. Achieving the "well-grooved"
movement of the free-throw or dart throw might also
occur through the Darwin Machine's process.
- Kicking and dancing might also benefit, to
the extent that the neural sequencer makes good
connections to output pathways for the lower body.
- Language would benefit, producing sequences of
words to make a sentence suitable to the occasion
and obeying the local rules of grammar as well.
- Plan-ahead in general might benefit, to the extent
that more general schemas could be sequenced
instead of movement commands.
- Insight, to the extent that it depends on testing
possible sequences of action by simulating them first
inside the head, ought to benefit from such
machinery and so allow a hominid to "do the right
thing" the first time without a lot of overt
trial-and-error.
- Musical abilities could similarly benefit from a more
general-purpose sequencer (stringing together chords
to make melodies) without any natural selection for
music per se.
- Serial-order recreations have surely been augmented
by secondary use of a sequencer, since it seems so
unlikely that chess and contract bridge have been
exposed to natural selection themselves. It is hard to
identify games that do not have a strong element of
either serial-order, ballistic movement, plan-ahead, or
fancy finger sequences.
Language Implications of a Neural Sequencer
One can imagine a manual-brachial sequencer being adapted
to simple kinds of language. A verb is usually a stand-in for a
movement. And the targets of a ballistic movement are examples
of nouns: "Throw at that rabbit!" or "Hit that nut." This seems
just like the predicate of a sentence: the verb and its object.
Spontaneous sign-language constructions in apes are often of this
type, as are the earliest utterances of human children.
All animals exhibit associative learning, but such sequencing
machinery might greatly augment associative abilities, quite without
making any initial use of the exact ordering. All humans go
through a stage of protolanguage (Bickerton, 1990) before the age
of two, where the mere association of words serves to carry a
simple message (and many a traveller reverts to protolanguage
when the foreign-language phrase book fails). Were chimpanzees
able to use combinations of their 36 standard vocalizations to
convey nonstandard messages, they could acquire some of the
power of pidgin languages of today.
Using a noun as the subject of a sentence-like construction
seems a bit more novel, with less precedent in movement planning
(where the subject is most often implied: oneself!). There is
nothing particularly verbal about such sentences: "You go there"
can be communicated with gestures, even eye movements. Having
two nouns in the same sentence may create some ambiguity as to
who is the actor and who is the acted-upon. If the
nouns are of very different classes (a person and a place), there
may be no ambiguity. Yet a construction such as "John called
Julie," where both nouns are capable of playing either role, starts
requiring syntax, such as the English convention that the word order
in a simple declarative sentence is actor-action-object. "Julie called
John" means something different than "John called Julie." The
mere addition of word order conventions would have enormously
expanded the number of possible messages that could be
constructed.
Bickerton (1990) cautions that much of modern language
requires more of a structural analysis than these examples of
sequencing provide. Beyond the second year of life, most normal
humans acquire an ability to construct sentences with lots of
grammatical words (below, before, because...) and
embedded phrases (He ate it with a spoon). Embedded
phrases tend to be nestled like Russian dolls rather than sequenced
like a string of beads, requiring what linguists call a phrase
structure analysis. And argument structure serves to identify the
thematic roles associated with a verb, as when "give" tends to
require a "Who", "What", and "to Whom." The verb "sleep"
merely sends one in search of a "Who" in the sentence.
To accomplish such modeling of the possible message, in a
manner far more complex than mere word-order allows, suggests
considerably more neural machinery for sequencing than two tracks.
Alternative interpretations of ambiguous sentences would need to be
maintained simultaneously for comparison against memories, and
better models evolved. It is this level of language which needs a
Darwin Machine in the brain, not simple association or word
ordering.
And so sequencing machinery could play a triple role: 1)
even a single sequencer augments the association of various
schemas, 2) it can also allow some conventions about word
order to be used to greatly expand the possible messages that
can be constructed, and 3) combinations of sequencers can permit
an evolution of mental models for "Who did what to whom?" that
use episodic memories of similar situations and sequences. Phrase
structure can be mimicked in a railroad-yard-like branching of
sequencer tracks; argument structure (and the other six theorems of
modern grammar) may simply be part of the judgements that shape
the evolving model. Any remaining ambiguity in the message (as
in Bickerton's example of The cow with the crumpled horn that
Farmer Giles likes) can often be resolved from memories
(perhaps Farmer Giles is known to collect horns, suggesting that it
is the horn that he likes, rather than the cow).
Besides the issues associated with innovative secondary
uses, we have the evolutionary issue of which function came first.
Or at least, which was fastest: Fast tracks in evolution tend to
preempt slow tracks. Even if the advantages of plan-ahead
intelligence or embedded phrases might suffice to evolve the more
elaborate neural sequencing machinery in the long run, might nut-cracking hammering have done it faster?
While we might agree that music ought to have so little
feedback on natural selection as to be near the bottom of the list,
the top of the list will surely be an interesting debate. Linguists
will be able to envisage the advantages of improvements in
representations and communications, and will suggest that language
improvements ought to be a fast track. Behaviorists will probably
favor problem-solving versatility as a fast evolutionary path. Since
I'm a neurophysiologist originally concentrating on the neurons that
make decisions about movements, my contribution to the debate
will be an evolutionary scenario for why accurate throwing might
be the fastest track of all.
Accurate Throwing as a Growth Industry
Many evolutionary improvements are "one-shot deals" that
cannot be repeated for further improvement; in a sense, they are
like college courses that cannot be repeated for additional credit.
Yet some, like the Music Department's performance courses, can be
taken again for credit, and so generate a significant growth curve.
For example: To an aquatic mammal with sufficient fat, less hair
seems to be better. And even less, better still. And so on until the
growth curve reaches a plateau (one can only become so naked).
What is the hominid growth curve for hammering nuts?
Given how skillful the chimpanzees studied by Boesch & Boesch
(1981) seem to be (at least, at positioning the targets; the
hammering sequence may not be as carefully varied), that curve
might have been close to a plateau even in our common ancestor.
Redoubling the profitability of nut-cracking probably wasn't done
very many times during hominid evolution, if our common ancestor
was chimplike in this aspect of behavior.
This is in contrast to throwing's growth curve. Chimpanzees
mostly engage in threat throwing, trying to intimidate leopards,
snakes, and fellow chimps; it seems to be an adjunct to flailing
about with a branch, rocks being secondarily used. The great
growth potential comes with converting the threat throw into a
predatory throw (see Plooij, 1978, for one possible scenario).
Action at a distance is an important invention for hunters, as it
reduces the chances of injury from hooves, horns, and teeth -- but
this is a good example of an improvement that cannot be repeated
for additional advantage.
What can be improved is the distance of the throw: twice
as far is always better, no matter how many times the distance
redoubles. And because throwing twice as far usually means
throwing twice as fast, the kinetic energy (or "stopping power" in
military terms) quadruples as distance doubles.
Accuracy can also be improved time and again for
additional advantage, as one comes to be able to hit smaller and
smaller targets at the same distance. The supply of small birds and
mammals is much more reliable than that of large mammals.
Attaining sufficient accuracy to strike the head is an important
improvement when dealing with medium-sized mammals.
Adding some technology to the rock throw further increases
the productivity of predatory throwing. The line of improvement
from spear to bow-and-arrow to crossbow is familiar to us, though
archaeology suggests that these were the products of the last ice age
and the present interglaciation; the line of improvement from the
common ancestor through Homo erectus to early Homo
sapiens has to be based largely on theory at this point.
A Throwing-based Evolutionary Scenario
In this exercise (which I hope that fans of tool-, language-,
or intelligence-based hominid evolution will consider emulating, so
as to facilitate comparisons), I will start with the skills that I will
ascribe to our common ancestor with the chimps and bonobos.
Thus I will assume that the basic ballistic skills were present (and
in the case of hammering, even well-developed). For throwing, we
don't know how accurate chimps really are; the anecdotes only
record their occasional (perhaps haphazard) hits. I think we can
assume that if chimps were accurate enough to bring down game
with any regularity, we would have heard about it (given how much
they enjoy fresh kills, they'd probably be the terror of Africa).
In evolution, changes in behavior tend to precede
adaptations in anatomy, so what kind of behavioral improvement
could a chimpanzee-like ape make that would be rewarded by a big
improvement in diet? Or in defense? If we assume that they
initially lack accuracy in throwing, this implies a big target or some
special circumstance. And the waterhole provides both.
Transition from Chimplike Behaviors. I suggest
that hominids practiced the familiar carnivore strategy: lying in
wait at the waterhole for herds to come to drink. Since
chimpanzees like to flail large tree branches around and then fling
them, assume one threw such a branch into the midst of a herd that
was busy drinking. Even before the branch landed, the herd would
startle from the sudden movement. By the time that the branch
landed somewhere in their midst, the herd would be wheeling
around and beginning to stampede.
Even if the branch didn't knock an animal down, some
animal might well become entangled with the branch and trip.
Ordinarily, such an animal would quickly get back on its feet and
successfully run away from the hunters. But in the midst of a
stampede, it would likely be knocked down again by other herd
animals in their panic. Some such animals are likely seriously
injured; even if the hominid had merely intended to chase away
competitors for scarce water, there might soon have been meat to
scavenge.
Once the hominids got the idea of flinging branches to gain
meat, the yield would have increased: the animal need not be
fatally injured by the branch or stampeding herd. It need only be
delayed enough in regaining its feet so that the hunters could
apprehend it. Certainly chimpanzees have no trouble dispatching
small pigs or monkeys, once they manage to lay hands upon them;
one imagines the hominids starting with some herd animal smaller
than zebra (Thomson's gazelles, for example) and tackling the
larger ones after their finishing-them-off technique improved.
I suggest that the waterhole fling technique survives to the
present day in the form of throwing sticks like the knobkerrie and
boomerang, used to trip up a running animal; its virtue as a
candidate for the earliest technique lies in its minimal requirements
for accuracy and its continuity with known abilities and preferences
of the chimpanzees. Flinging branches into herds at a waterhole
would have made a good transition onto the bottom of the ladder of
predatory throwing; the herd not only makes a large target (it
doesn't matter which one is hit) but the herd "amplifies" the effect
of the projectile by delay or injury of a downed animal. And the
technique would be well-rewarded, what with a new-found ability
to eat meat every day; one can imagine a "new niche" level of
boom times resulting for the hominids that invented this technique.
Transition to Rock Throwing: Lacking a handy
branch (and the technique might well exhaust the supply near a
waterhole), chimps tend to throw a big rock. Thrown into a herd, it
might well land between animals (and so be less effective than a
large branch). But herds that are feeling threatened by predators
tend to cluster together tightly, and this is especially notable upon
visits to the waterhole. Even if you miss, it's easier next time!
A large rock landing on the back or side of a small grazing
animal might well injure it sufficiently; however, all the rock really
has to do is knock the animal off its feet and the stampeding herd
will serve to delay its escape. For larger herd animals, however,
one can imagine a rock as being frequently ineffective, except on
the rare occasions when an animal is hit on the head. What might
improve the technique?
Enter the Handaxe: A flat rock (a chunk of slate,
for example) may spin in flight like a Frisbee. If thrown in the
plane of spin, air resistance is minimized and distance increased. If
it hits an animal this way, it may also do more damage, as all of
the impact is concentrated in a narrow edge; the sharper the edge,
the more damage to the skin.
By itself, this does not seem very promising. I mention it
because it seems to describe the enigmatic Acheulean handaxe in
several respects: nice spin symmetry, and sharpened edges all
around. Those sharpened edges are the prime reason that
"handaxe" is surely a misnomer: pounding with one would cause it
to bite the hand that held it. While damaged versions of the classic
shape might well serve a chopping function, surely the symmetry,
the edging, and the point are indicative of some function that we
have failed to comprehend. I suggest that classic handaxes may be
better appreciated in the context of the waterhole ambush.
Following the suggestion of Jeffreys (1965) that handaxes
might have been thrown with spin, e.g., into a flock of birds,
O'Brien (1981, 1984) undertook to test the aerodynamic properties
of a fiberglass replica of one of the very large Acheulean handaxes
from Olorgesaile. Even when thrown by experienced athletes, the
handaxe did exactly what Frisbees do when thrown by beginners:
it turned from horizontal to vertical orientation in mid-flight and
then landed on edge. I recently repeated her experiments with five
heavily-weathered handaxes from the Sahara, with much the same
results. Unlike Frisbees that roll along the ground for some
distance, a spinning handaxe will soon impale its point in the
ground and come to a sudden halt.
While of no significance when throwing into a flock of
birds, I can see distinct advantages of such properties when lobbing
a handaxe into a herd visiting a waterhole. An ordinary rock,
lobbed into a tightly-packed herd, is most often going to hit an
animal on its back or side. Generally the rock will then bounce off
-- but if it were to somehow impale itself, it would transfer all of
its momentum to the animal, rather than only a fraction.
Consider throwing a handaxe into the herd. It lands on
edge, doing considerable damage to some animal's skin (whether or
not the edge actually penetrates it). The handaxe spins forward and
the point tends to snag the skin (catching a roll of skin pushed
forward by the impact, even if it doesn't penetrate). Then the
handaxe drops to the ground, its forward momentum exhausted
since it was effectively transferred to the animal's back or side.
This is far more likely to knock the animal off its feet than is the
impact of an ordinary rock that bounces away.
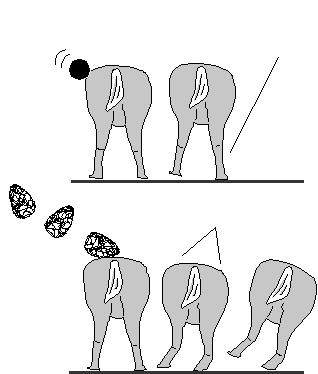 Neurological Knock-down: While the
foregoing might suffice, I have discovered (Calvin, 1990) some
additional neurological reasons why even a small handaxe (and
most will fit comfortably into the hand) might bring down a large
animal: the skin damage tends to countermand the reflex
commands that ordinarily would prevent toppling after impact.
Neurological Knock-down: While the
foregoing might suffice, I have discovered (Calvin, 1990) some
additional neurological reasons why even a small handaxe (and
most will fit comfortably into the hand) might bring down a large
animal: the skin damage tends to countermand the reflex
commands that ordinarily would prevent toppling after impact.
As background, note that all mammals have righting
reflexes; a blow pushing the animal to its right will tend to generate
extension commands to the right legs. If the response is quick
enough, the animal's slow rightward movement from the momentum
transfer will be countered and the animal will manage to keep its
center of gravity to the left of its right legs. Once the center of
gravity passes the supports, the animal will collapse (the prime
virtue of higher velocities and bigger rocks is that the rightward
drift becomes so rapid that instability is reached before the reflex
loop can operate).
But there are other reflexes which may overwhelm righting
reflexes, e.g., the withdrawal reflexes which tend to create
movement away from a site of injury, as when one lifts the leg
after stepping on something sharp. Withdrawal reflexes depend on
the site of injury; for a four-legged animal, injury to its back
(which would most often come from an overhanging tree branch or
rock outcrop) causes both hindlegs to flex, dropping the back down
to disengage it from the overhang. Pain is the effective stimulus
for a withdrawal reflex, and the handaxe's sharpened edge would
surely cause much more pain than the featureless rock. Still, there
are many examples (see Calvin, 1983a) of people not noticing skin
cuts if busy with something else.
It is manipulation of the edges of an incision which cause
the most requests from patients for boosters in the local anesthetic.
A handaxe that injured the skin, and then yanked hard on the
damaged area (as would occur when the handaxe's point snagged
the skin), seems likely to cause a massive flexion reflex of the
hindlimbs. This alone might cause the animal to collapse, even if
the sideward momentum transfer was minimal because it was a
small handaxe.
So the point need not penetrate, nor need the edges incise,
the skin for such a use to be effective. Nor need the handaxe be
heavy, or have a lot of horizontal velocity, as pushing the animal
sideways is no longer the only way to make it collapse. (Needless
to say, the hunter need not understand neurological knock-down
mechanisms in order to have noticed that even a juvenile's small
handaxe was effective, so long as it had a sharp edge and point).
Still, this knock-down might merely result in a limping animal
running away from the pursuing hunters -- were it not for the herd
stampeding and delaying the downed animal. The context of the
herd is still essential (which also means that the technique would be
ineffective against fellow hominids -- unless they clustered together
for protection), at least until the side-of-the-barn accuracy improves.
I suggest that the handaxe's characteristic features (flattened
spin symmetry, all edges sharpened, and something of a point)
make it an excellent projectile for waterhole predation, one that
might have greatly increased the yield. That handaxes (Howell,
1961, Isaac, 1977, Kleindienst & Keller, 1976) are regularly found
along watercourses and lake margins (as if lost in the mud like a
golf ball; often they are standing on edge in situ) supports this
explanation. While handaxes also make good fleshers (and broken
ones may suffice as choppers), none of the other uses proposed for
handaxes can explain all of the afore-mentioned characteristic
features or the archaeological peculiarities. I would suggest that the
Acheulean cleavers, and the handaxes without all-around edging,
may be damaged versions of the classic design; if they were
thought to be less effective in downing herd animals, they might
well have been "retired" and converted to less demanding secondary
uses such as chopping or fleshing. The handaxes found in lake
beds are probably the ones that the hunter lost in the mud (Indian
arrowheads in North America, as well as golf balls, similarly
accumulate at waterholes). Thus the theoretical prediction:
waterhole handaxes ought to be more completely edged, in
comparison to the ones found elsewhere.
Improvements Beyond the Handaxe Lob: The
waterhole lob ought to work well on the savannah, with its herds
and obligate waterholes during the dry season that must be visited,
predators or not. While flinging branches into such herds suggests
a natural transition from chimpanzee-like behaviors, and while the
handaxe might have been the "high technology" of this waterhole
ambush technique, we must ask where the growth curve goes from
there. There are some important inventions, such as persuading
animals to injure themselves by stampeding over a cliff, but their
growth curves are minor.
Longer and longer lobs would be useful as herds became
wary, but the real growth curve lies with accuracy: being able to
hit even a small herd. Or being able to prey on animals that do not
cluster together so conveniently and compound the injury done the
downed animal. Or being able to prey on the smaller mammals and
birds which are far more plentiful (and which, when their numbers
are depleted by heavy predation, can regain population numbers
more quickly because of their shorter gestation times and juvenile
periods).
The only problem is that accuracy is expensive, whether
trying to hit a smaller target from the same distance or trying to hit
the same target from twice the distance. This is because of one of
the fundamental differences between the electrical operation of our
excitable cells (both nerve and muscle) and that of modern
electronics; because of the way that ion channels through the cell
membrane function, the cells that control our movements cannot
time events with any great accuracy. They are jittery, irreducibly
noisy.
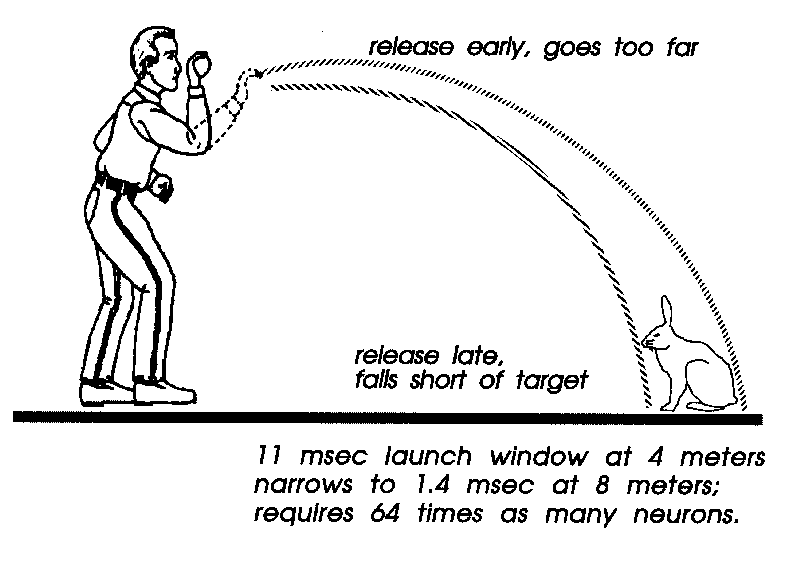 And throwing has a crucial timing step that is not present in
hammering or clubbing or kicking: the projectile must be released
at just the right instant (Fig. x.2). Too early, and the launch angle
will be too high, the projectile overshooting the target. Too late,
and the projectile hits the ground in front of the target. We can
talk of the "launch window" as that span of release times where the
projectile will hit the target somewhere between its top and bottom.
For a throw at a rabbit-sized target from a four meter distance
(about the length of an automobile), this launch window is 11 ms
wide. So at the end of a throw which started several hundred ms
earlier, one must time the relaxation of the grip to stay within that
11 ms. The typical spinal motorneuron has intrinsic timing jitter of
at least that much (Calvin & Stevens, 1968; Calvin, 1983b, 1986),
when the cell is generating action potentials at 200 ms intervals.
And throwing has a crucial timing step that is not present in
hammering or clubbing or kicking: the projectile must be released
at just the right instant (Fig. x.2). Too early, and the launch angle
will be too high, the projectile overshooting the target. Too late,
and the projectile hits the ground in front of the target. We can
talk of the "launch window" as that span of release times where the
projectile will hit the target somewhere between its top and bottom.
For a throw at a rabbit-sized target from a four meter distance
(about the length of an automobile), this launch window is 11 ms
wide. So at the end of a throw which started several hundred ms
earlier, one must time the relaxation of the grip to stay within that
11 ms. The typical spinal motorneuron has intrinsic timing jitter of
at least that much (Calvin & Stevens, 1968; Calvin, 1983b, 1986),
when the cell is generating action potentials at 200 ms intervals.
But most people can, with practice, hit a rabbit-sized target
at such a comfortable distance on perhaps half of the tries, so let us
assume that the timing mechanisms are sufficient. Move the target
out to twice the distance, eight meters, and practice enough to
achieve hits on half the tries. The reason that this is so hard is that
the launch window shrinks a factor of eight to only 1.4 ms. The
solid angle subtended by the target fell by a factor of four;
furthermore, throwing twice as far with a reasonably flat trajectory
means throwing twice as fast, so the time scale halved. An
electronic instrument would have no trouble keeping its timing jitter
to within a 1.4 ms window over several hundred milliseconds. But
neurons would probably have to be redesigned from scratch.
The Law of Large Numbers. Nature seldom
redesigns, but it does one thing very well: it duplicates cells
endlessly. And it seems that the way around cellular jitter is to
simply assign a great many neurons to the same task, timing the
launch, and then averaging their recommendations. Need to halve
the jitter? Just use four times as many cells as originally sufficed.
To double the throwing distance and maintain hit rate, one needs an
eight-fold reduction in jitter: that merely requires 64 times as many
timing cells as originally sufficed. Triple the distance? Only 729
times as many cells.
Now getting good enough to re-attain your hit rate at twice
the distance is surely not a matter of growing a lot of new cells for
the movement control circuits. It seems more likely that they are
borrowed as one gets set to throw, the experts (probably the
premotor cortex and the cerebellum) recruiting some temporary help
from other brain regions, much as the choir in the Hallelujah
Chorus borrows the audience temporarily.
Might, however, our four-fold increase in brain size have
something to do with this Law of Large Numbers principle, our
developmental genes growing more nerve cells as a result of
throwing accuracy coming under natural selection for feeding
families during hominid evolution? Alas, a four-fold increase in
neuron numbers will only "buy" a 26 percent increment in throwing
distance -- and the first 10 percent increase in neuron numbers
(about what the natural variation in the human population might
presently be) will only buy a 2 percent farther throw. And so I
doubt that it was a simple matter of bigger-brains-throw-better; I
suspect that recruiting the helpers is easier with a more juvenilized
brain (Calvin, 1990), and that the bigger brain is simply a side-effect
of neoteny, that there was not selection for size per se.
Concluding Remarks
Many things have contributed to hominid evolution, and
humans wouldn't be what we are today without effects from many
of them. The inventions that would seem to make the most
demands on brain reorganization, however, are accurate throwing
and structured language.
Predatory throwing seems possible from such transition
stages as my waterhole lobbing proposal; accurate throwing is not a
single invention but a long climb, fortunately with a growth curve
that seems unlimited. Eating meat regularly would have greatly
expanded the hominid niche, allowing hominids to expand well
outside the savannah, even into the temperate zone (where the
major constraint is that the gathering is so poor in the winter
months that either hoarding or meat-eating seems essential).
Throwing seems to be a fast track for hominid evolution;
whether it is the fastest track to a versatile sequencer and its
Darwin Machine secondary uses can be better judged when a
similar analysis is available for language, intelligence, toolmaking,
and other candidates. What are their transitions from apelike
behaviors, their growth curves, their niche-expanding properties,
their secondary uses?
ACKNOWLEDGEMENTS
I thank Derek Bickerton, Kathleen Gibson, and Katherine
Graubard for comments on the manuscript; revision of the
manuscript was greatly aided by conversations with Christopher
Boehm, Christophe Boesch, David Brin, Iain Davidson, Daniel C.
Dennett, Dean Falk, Susan Goldin-Meadow, Doug Hofstadter, Nick
Humphrey, Ray Jackendoff, Adam Kendon, Marcel Kinsbourne,
Philip Lieberman, Andy Lock, Peter MacNeilage, William McGrew,
George Ojemann, Martin Pickford, Duane Rumbaugh, Vince Sarich,
Sue Savage-Rumbaugh, Mark Sullivan, Nick Toth, Peter Warshall,
Pete Wheeler, and Thomas Wynn.
Fig. x.1.
The basic design and operation of a Darwin Machine: