 Webbed Reprint Collection
Webbed Reprint Collection
William H. Calvin
University of Washington
Box 351800
Seattle WA 98195-1800 USA
Email || Home
Page || publication list
|
William H.
Calvin
"Cortical Columns, Modules, and Hebbian
Cell Assemblies"
as it appeared in
The Handbook of Brain Theory and Neural Networks,
edited by Michael A. Arbib (Bradford Books/MIT Press), pp.269-272
(1995).
copyright ©1995 by William
H. Calvin and MIT Press
|
1. INTRODUCTION
Our cerebral cortex sits atop the white matter like a
thin layer of icing, about 2 mm thick. About 148,000
neurons lay under each square millimeter; however, it is
this "icing" that is horizontally layered, not the "cake"
beneath. Neurons with similar interests tend to be
vertically arrayed in the cortex, forming cylinders known
as cortical columns (though they are sometimes
elongated curtainlike bands); minicolumns. are
about 30 um in diameter while macrocolumns are
0.4-1.0 mm.
The superficial cortical layers of the primary
visual cortex in primates (V1) have regions that contain
neurons particularly sensitive to color; these "blobs"
selectively project to color-sensitive zones in area V2
called "stripes." The "blobs" are separated by
macrocolumnar distances (Bartfeld & Grinvald, 1992),
with surrounding regions containing neurons more
sensitive to visual form than to color; however, only 30-35 percent of blob neurons are color-sensitive and many
animals with poor color vision nonetheless have blobs.
Besides the color stripes of V2, there are stripes in
parallel which specialize in form, others in binocular
aspects (Livingstone & Hubel, 1988).
Columns, barrels, blobs, and stripes have all
been called cortical modules, and the term is
frequently applied to any segmentation or repeated
patchiness (Purves et al, 1992). By so loose a definition,
both a dried-up river bed and the fur of a "marmalade"
cat are also modular. Frequently there is no evidence of
either function or detailed internal circuitry, just
premature talk of "repeating patterns of circuitry" and
"iterated modular units." Module is also loosely
used by theoreticians to mean any functional grouping
in the brain, also violating the notion of repeating
standardized units. Some columns may indeed be
modular by the usual conventions of modular furniture
and modular electronics, but a favorite candidate for a
higher-order module, the "hypercolumn," has now been
shown not to be modular at all (Blasdel, 1992).
Why should neurons with similar functional
specializations cluster together? Why should some
clusters extend vertically through most of the cortical
layers? The reasons could be functional (Bullock, 1980;
Shaw et al, 1982) or merely an epiphenomenon of
development (Purves et al, 1992). Here the problem will
be approached via clustering tendencies provided by the
cortex's horizontal connections (Katz & Callaway, 1992).
1.1 Layering and its Functional Correlates
Traditionally six neocortical layers were
distinguished by neuroanatomists, but this has been
subject to some lumping and splitting. Layers II
and III can usually be lumped together, as when
one talks of the superficial pyramidal neurons. But layer
IV has had to be repeatedly subdivided in the
visual cortex (IVa, IVb, IVcAlpha,
IVcBeta).
Layer IV neurons send most of their
outputs up to II and III. Some
superficial neurons send messages down to V
and VI, though most connect (either laterally or
via U-fibers in white matter) in their own layers. Layer
VI sends messages back down to the thalamus
via the white matter while V sends signals to
other deep and distant neural structures, sometimes even
the spinal cord. A simple path would come into
IV, then up to III, down to V or
VI, and then back out of the cortex to some
"subcortical" structure (White, 1989). So for any column
of cortex, the bottom layers are like a subcortical
OUT box, the middle layer like an IN
box, and the superficial layers somewhat like an
INTEROFFICE box connecting the columns and
different cortical areas. These are not, of course,
exclusive roles, e.g., VI pyramids also have axon
branches extending horizontally to terminate in
IV.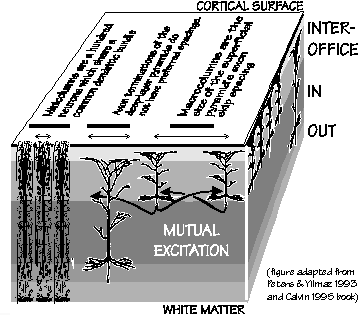
Traditionally, the association cortex was simply
terra incognita, the 90% of human cerebral cortex
that isn't motor strip or a primary sensory area.
However, Diamond (1979) argues that the "motor cortex"
isn't restricted to the motor strip but is the fifth layer of
the entire cerebral cortex. That's because V,
whatever the area, contains neurons that at some stage
of development send their outputs down to the spinal
cord, with copies to the brain stem, basal ganglia, and
hypothalamus. Diamond likewise argues that the fourth
layer everywhere is the "sensory cortex" and that second
and third layers everywhere are the true "association
cortex." Calvin (1994) argues that the superficial layers
have the right properties to implement darwinian
processes on the time scale of thought and action.
1.2 Columnar Clustering
A given neuron, however, may have dendrites
spanning a few layers, especially if it is a pyramidal
neuron: these taproot-shaped cells are the excitatory
neurons of neocortex. The other neuron types are
thought to be inhibitory (exception: IV's spiny
stellate neuron is excitatory). In Nissl stains that show
only cell bodies of neurons, Ramon y Cajal saw narrow
columns (hereafter, minicolumns) running from
white matter to the cortical surface; the cell-sparse gaps
are about 30 um apart in human cortex. It now appears
that the gaps are vertical bundles of axons and apical
dendrites. In monkeys, these minicolumns are 31 um
diameter but in cats they are about twice that diameter
(Peters & Yilmaz, 1993). In monkey, there are about 100
neurons in such a minicolumn (143 in V1), 39 of which
are superficial pyramidal neurons.
The human cerebral cortex totals 2,200 cm2;
unfolded, it would fit on four sheets of letter-sized
paper. That of the apes would fit on a single sheet, that
of cats and monkeys on a postcard, and that of a rat on
a small postage stamp. On the basis of layering
differences, there are 52 "areas" in each human
hemisphere; a Brodmann area averages 21 cm2 and 250
million neurons grouped into several million
minicolumns. Physiologically, there are aggregations
(hereafter, macrocolumns) which would seem to
contain, at most, a few hundred minicolumns; this may
be secondary to an organization of the input wiring into
projection macrocolumns (see Figure).
In 1957, Mountcastle and coworkers discovered
a tendency for somatosensory strip neurons responsive
to skin stimulation (hair, light touch) to alternate with
those specializing in joint and muscle receptors about
every 0.5 mm. It now appears that there is a mosaic
organization of similar dimensions, the neurons within
each macrocolumn (or "segregate") having a receptive
field optimized for the same patch of body surface (see
Favorov & Kelly, Somatosensory system).
Hubel and Wiesel, recording in monkey visual cortex,
saw curtainlike clusters ("ocular dominance columns")
which specialized in the left eye, with an adjacent cluster
about 0.4 mm away specializing in the right eye.
Orientation columns are of minicolumn
dimensions, within which the neurons prefer lines and
edges that are tilted about the same angle from the
vertical; there are many such minicolumns specializing
in various angles within an ocular dominance
macrocolumn (Hubel & Wiesel, 1977). The visual cortex
provides us with our best insights into cortical circuitry
(see Lund et al, Visual cortex). The relationships
between minicolumns and macrocolumns are best seen
there (see Miller, Ocular dominance and orientation
columns), though it is hazardous to generalize
because ocular dominance columns themselves are less
than universal, e.g., they are not a typical feature of New
World monkeys.
1.3 Horizontal Organization in Neocortex
Lateral connections in an array are similar to
the relaxation algorithms used for neurallike problem-solving.
Recurrent inhibition is thought, in cortex
as elsewhere, to provide an antagonistic organization
that sharpens responsiveness to an area far smaller than
would be predicted from the anatomical funneling of
inputs (see Barlow, Lateral inhibition).
Recurrent excitation may be especially prominent
in the superficial layers of primate neocortex. The
superficial pyramids send myelinated axons out of the
cortical layers into the white matter; their eventual
targets are typically the superficial layers of other
cortical areas when of the "feedback" type; when
"feedforward" they terminate in IV and deep
III. Long corticocortical terminations are often
organized into interdigitating macrocolumns.
But superficial pyramidal neurons also send
unmyelinated collaterals to adjacent superficial
pyramids: roughly 70 percent of the excitatory synapses
on any superficial pyramid (but less than 1 percent of
those on layer V pyramids) are derived from
pyramidal neurons less than 0.3 mm away. In general,
the average cortical neuron contacts fewer than 10
percent of all the neurons in that radius, so superficial
pyramids may be said to have an unusually strong
propensity to excite one another.
There is also an unusual patterning to such
superficial connections that suggests a columnar
organizing principle. The collateral axon travels a
characteristic lateral distance without giving off any
terminal branches; then it produces a tight terminal
cluster (see Fig. 5 in Gilbert, 1993). The distance to the
center of the terminal patch is about 0.43 mm in primary
visual cortex, 0.65 mm in the secondary visual areas, 0.73
mm in sensory strip, and 0.85 mm in motor cortex of
monkeys (Lund et al, 1993). The axon may then
continue for an identical distance and then produce
another cluster, and this occasionally continues for some
mm.
Because of this local standard for axon length,
recurrent excitation becomes probable among some cell
pairs. Horizontal connections are also found among the
pyramidal neurons of V and VI, but the
regular spacing has been noted only for the pyramids of
the superficial layers. In the absence of simultaneous
recordings at appropriate spacings, we can only guess at
the consequences of the mutual re-excitation. Even if the
synaptic strengths were high, much longer axons would
be required to give conduction times that escape
refractoriness and create (even briefly) the proverbial
reverberating circuit (corticocortical conduction velocities
are about 0.3-0.5 mm/msec).
1.4 Synchrony-shaped Connectivity
Recurrent excitation, however, can induce
synchrony: even weak coupling between relaxation
oscillators is known to quickly produce entrainment (see
Segundo, Stiber, & Vibert, Entrainment and
synaptic coding of spike trains). Were the cells
otherwise active, they would soon tend to produce some
spikes at about the same time. Cells in cortical
minicolumns often fire in synchrony; more widespread
synchronized firing has been a recent theme in cortical
neurophysiology; such synchrony occurs more frequently
during difficult tasks and is the best-known correlate of
perceptual binding (see Singer, Synchronization
of neuronal responses as a putative binding
mechanism).
Synchronized firing in the context of the
superficial layers of neocortex has some important
implications for synaptic enhancement. The local field
potentials associated with synchronized firing have their
source in the superficial layers. The long-term
potentiation (LTP; it is dependent on sufficient
postsynaptic depolarization by other inputs)
demonstrated in motor cortex is confined to those layers,
and that is where most N-methyl-d-aspartate
(NMDA) receptors for glutamate are located.
In such a system, near-simultaneous arrivals
may enhance the synaptic strength of the coincident
inputs -- but, even more importantly, repeated
coincidences should be particularly effective in NMDA-like synapses. One can imagine synchronous "test
patterns" during development serving to shape up adult
cortical connectivity via use dependence and selective
survival. The superficial pyramid's lateral connections
suggest that they could organize synchronous clusters
about 0.4 mm apart in primary visual cortex -- which
happens to be the approximate size of the macrocolumns
of Area 17, the ocular dominance columns. It is thought
that ocular dominance columns are organized by a
gradual segregation of geniculate afferents into layer
IV, but such selective survival might be
secondary to neural activity during development and
thus dependent on connectivity in the superficial layers.
2. DISCUSSION
2.1 Permanent and Temporary Macrocolumns
Given that both are about a half millimeter, what is
the relationship between the permanent macrocolumns
and the ephemeral entrained pairs, whose synchrony can
be destroyed in an instant by a wave of inhibition?
Entrained cells probably form triangular
mosaics on occasion, given that a superficial pyramid
sends axon collaterals in many directions. Two
entrained cells may send axon terminals to an
equidistant point, with impulses arriving simultaneously,
and so entrain it as well. This equilateral triangle forms
the basis of a triangular mosaic; the spacing will be the
local metric, whether 0.43 mm or 0.85 mm (hereafter,
simply "0.5 mm"). Because the basis of recruitment and
entrainment is conduction time, not distance per se,
various distortions of the triangular mosaic might be
seen if conduction velocity (typically 0.3-0.5 mm/msec
within cortex) and synaptic delay are not constant.
Just from the geometry, the overall impression
should be like that of wallpaper, where corresponding
points in a repeating pattern can be identified -- but
where the "origin" of the pattern cannot necessarily be
identified. One can, however, artificially inscribe a
boundary such that homologous points are approached
but not included. If one does this with many
neighboring points, each of which is part of a different
triangular mosaic, the largest possible "unit area"
without repetitions will be hexagonal in shape. The
distance between parallel sides of the hexagon will be 0.5
mm as well. Like the wallpaper's unit pattern, this
hexagon need not have a unique "origin" and it might be
meaningless to speak of a hexagonal boundary unless
underlying resonances with synaptic strengths were so
organized. One could speak (Calvin & Ojemann 1994)
however, of this hexagonal unit pattern being laterally
copied or cloned, thanks to the triangular mosaic
tendencies of recurrent excitation among superficial
pyramids.
Because a pair of synchronized neurons could
form a triangular mosaic that is not parallel to the
others, hexagonal mosaic cloning depends, in effect, on
two identical and adjacent hexagons of spatiotemporal
patterning. This parent pair could be due to evoked
activity in an input pathway, or the initial
spatiotemporal pattern could be generated by cortical
connectivity just as the gaits of locomotion are generated
by spinal cord circuitry. The spatial-only connectivity
patterns in cortex that generate the spatiotemporal firing
patterns could themselves be contained in adjacent
hexagons; were this visible to anatomical or
physiological techniques, we might consider it a
permanent macrocolumn. Otherwise, we would merely
see transient physiological ensembles of macrocolumnar
dimensions with hexagonal repeats.
2.2 Modular Aspects
It is widely expected that cerebral cortex will
turn out to have circuits which, in different cortical
patches, are merely repeats of a standard pattern.
However, module has instead been loosely
applied to a wide range of functional or anatomical
collectives. A true module would be a cortical patch
which, whatever the origin of its inputs or the
destination of its outputs, nonetheless has an internal
architecture which is the same from one instance to
another, with only minor differences in local tuning.
The best candidate for a true module was the
"hypercolumn" (Hubel & Wiesel, 1977): two adjacent
ocular dominance columns, each containing a full set of
orientation columns. Adjacent hypercolumns would
merely represent different patches of the visual field;
inputs and outputs might differ, but internal wiring
should be similar from one hypercolumn to the next.
However, newer mapping techniques have
shown that ocular dominance repeats are somewhat
independent of orientation column repeats (Blasdel,
1992), making adjacent hypercolumns internally
nonidentical, i.e., not iterated. For this reason,
"hypercolumn" now appears with scare quotes around it
in the visual system literature. No current use of
"cortical module" bears any relationship to the
technological use of the term; cortical
cluster would be a more appropriate term.
2.3 The Hebbian Cell Assembly
Representations via a pattern are familiar from
the trichromatic theory of color perception and the
similar aspects of taste. Evoking the memory of your
grandmother's face is probably not a matter of activating
a single specialized neuron; rather, it is thought to
involve the activity pattern in an ensemble of cortical
neurons, each of which helps to implement other
memories as well. Different sensory or motor schemas
might be characterized by different firing patterns in
time and space, each characteristic spatiotemporal
pattern serving as a code for an item from an
individual's memory.
Memory recall may consist of the creation of a
spatiotemporal sequence of neuron firings, probably one
similar to that present at the time of the input to
memory, just shorn of some of the nonessential frills that
promoted it. It would be like a message board in a
stadium, with lots of little lights flashing on and off, but
creating a pattern. A somewhat more general version of
a Hebbian cell assembly (Calvin & Ojemann 1994)
would avoid anchoring the spatiotemporal pattern in
particular cells, to make it more like the way the
message board can scroll. The pattern continues to
mean the same thing, even though it's implemented by
different lights. Though we tend to focus on the lights
which turn on, note that lights which remain off also
contribute to the pattern.
The notion of convergence zones for associative
memories raises the issue of maintaining the identity of
a spatiotemporal code during long-distance
corticocortical transmission, such as through the corpus
callosum. Distortions of the spatiotemporal pattern by a
lack of precise topographic mappings might be
unimportant where the information flows in only the
one direction. But because the connections between
distant cortical regions are typically reciprocal, any
distortions of the original spatiotemporal firing pattern
during forwards transmission would need to be
compensated in the reverse path in order to maintain the
characteristic spatiotemporal pattern as the local code for
a sensory or motor schema. In addition to inverse
transforms or error-correction mechanisms, degenerate
codes seem possible (as when six different RNA triplets
all code for leucine).
Error correction is particularly interesting
because reliable copying capabilities often provide
insight into possible codes, e.g., the genetic code puzzle
was solved after identifying which physical patterns
could be readily duplicated. Are there physiological
clones of cerebral schemas that might help us identify a
cerebral code such as the relevant Hebbian cell
assembly?
Spatiotemporal pattern copying was inferred
from the need to reduce timing jitter during precision
throwing using Law of Large Numbers averaging. The
triangular mosaic of electrical activity predicted from
standard-length axons has an inherent error-correction
property that can maintain spatiotemporal patterns
during copying, so long as they are small enough to fit
inside a macrocolumn-sized hexagon of cerebral cortex.
This suggests that hexagonal Hebbian cell assemblies
could implement a cerebral code (Calvin, 1994).
References
* especially suitable for getting oriented
Bartfeld, E., and Grinvald, A., 1992, Relationships
between orientation-preference pinwheels, cytochrome
oxidase blobs, and ocular-dominance columns in
primate striate cortex. Proc. Natl. Acad. Sci. USA
89:11905-11909.
*Blasdel, G.G., 1992, Orientation selectivity,
preference, and continuity in monkey striate
cortex. J. Neurosci. 12:3139-3161.
Bullock, T.H., 1980, Reassessment of neural
connectivity and its specification, in Information
Processing in the Nervous System, (H.M. Pinsker
and W.D. Willis, Jr., Eds.), New York: Raven
Press, pp.199-220.
Calvin, W.H., 1994, The emergence of intelligence.
Scientific American 271(4):100-107.
Calvin, W.H., Ojemann, G.A., 1994, Conversations with Neil's Brain:
The Neural Nature of Thought and Language. Reading MA: Addison-Wesley.
Diamond, I., 1979. The subdivisions of neocortex:
A proposal to revise the traditional view of
sensory, motor, and association areas, in Progress
in Psychobiology and Physiological Psychology 8
(J.M. Sprague, A.N. Epstein, Eds.), New York:
Academic Press, pp. 1-43.
Gilbert, C.D., 1993, Circuitry, architecture, and
functional dynamics of visual cortex. Cerebral
Cortex 3:373-386.
Goldman-Rakic, P., 1990. Parallel systems in the
cerebral cortex: the topography of cognition, in
Natural and Artificial Parallel Computation (M.A.
Arbib, J.A. Robinson, Eds.), Cambridge: M.I.T.
Press, pp.155-176.
Hubel, D.H., Wiesel, T.N., 1977, Functional
architecture of macaque visual cortex. Proc. Roy.
Soc. (London) 198B:1-59.
Katz, L.C., Callaway, E.M., 1992, Development of
local circuits in mammalian visual cortex. Ann.
Rev. Neurosci. 15:31-56.
*Livingstone, M.S., Hubel, D.H., 1988, Segregation
of form, color, movement, and depth: Anatomy,
physiology, and perception. Science 240:740-749.
Lund, J.S., Yoshioka, T., Levitt, J.B., 1993,
Comparison of intrinsic connectivity in different
areas of macaque monkey cerebral cortex.
Cerebral Cortex 3:148-162.
*Mountcastle, V.B., 1979, An organizing principle
for cerebral function: The unit module and the
distributed system, in The Neurosciences Fourth
Study Program (F.O. Schmitt and F.G. Worden,
Eds.). Cambridge: MIT Press, pp.21-42.
Favorov, O.V., Kelly, D.G., 1994, Minicolumnar
organization within somatosensory cortical
segregates: I. Development of afferent
connections. Cerebral Cortex 4:408-427.
Peters, A., Yilmaz, E., 1993, Neuronal organization
in area 17 of cat visual cortex. Cerebral Cortex
3:49-68.
*Purves, D., Riddle, D.R., LaMantia, A-S., 1992.
Iterated patterns of brain circuitry (or how the
cortex gets its spots). Trends in the Neurosciences
15:362-368 (see letters at 16:178-181).
Shaw, G.L., Harth, E., Scheibel, A.B., 1982,
Cooperativity in brain function: assemblies of
approximately 30 neurons. Exp. Neurol. 77:324-358.
White, E. L., 1989. Cortical circuits Boston:
Birkhauser.
The figure is NOT the exact one which appears in the handbook
but an improved one which appears in W. H. Calvin, The Cerebral Code (MIT Press 1996).
mail@williamcalvin.com
|| Home Page || Calvin
publication list || My Science Surf
column || The
Calvin Bookshelf || Revised 5 December 1996
