William H. Calvin and George A. Ojemann, Inside the Brain: Mapping the Cortex, Exploring the Neuron (New American Library, 1980), chapter 11. See also http://WilliamCalvin.com/ Bk1/bk1ch11.htm. |
William H. Calvin
University of Washington |
|
11 Circuits for Seeing: |
©1980 by William H. Calvin and George A. Ojemann |
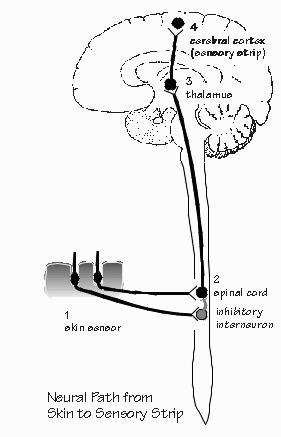
The screen goes blank after Neil finishes with the last slide. Immediately, everyone in the OR moves around a little and the noise level rises, as all had been quiet during the performance and recording. "Very nice, Neil. Want to move around a little?" asks the neurosurgeon. "Aagh! My leg fell asleep. Now my toes are tingling. Is that important? I wouldn't mention it, but you told me to tell you if I felt something," says Neil. "Sounds familiar. That happens to nerves when they get blocked by pressure. No problem as long as they're tingling and not numb. The anesthesiologist will check out things. just move around a little." There is a chain of neurons which links the skin with the sensory strip of Neil's cerebral cortex. First is the transducer neuron in the skin, which senses pressure, the movement of a hair, or temperature. Next comes the "second-order" neuron, across the synapse in the spinal cord or in the brain stem. The third neuron in the chain is often in the thalamus. The axons from the thalamus go up to the sensory strip of cerebral cortex, so that there arc at least three neurons in the chain before a cortical cell gets a chance. The direct four-neuron chain is not, however, the only path. As we have seen, there may be a relay in the brain stem's reticular activating system. Some axon branches go up to the cerebellum, the elaborate infolded structure which sits atop the brain stem and assists in the coordination of movements and posture. It too talks to the sensory strip. Each of the senses has a similar set of direct and indirect pathways from the transducers to the cerebral cortex.
It is customary to think of this as a relay race, with something simply being passed on from one neuron to another. It is, however, more analogous to the spread of a rumor, As each successive neuron retells the story, It may leave out certain details, add others, or perhaps not retell the story at all. Each neuron has its own characteristic style of storytelling.
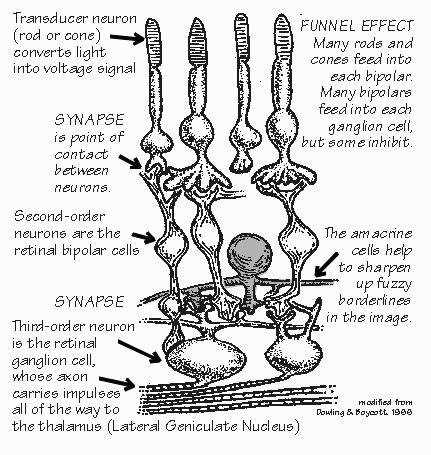
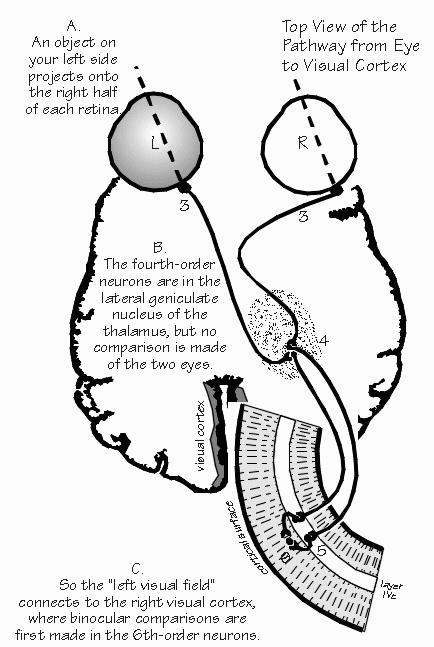
Particularly in the case of vision, neurophysiologists have been able to partially understand what happens at each successive neuron in the chain. The transducer neurons are the rods and cones of the retina. The second and third neurons of the chain are also in the retina. The axon of the third neuron, called the retinal ganglion cell, runs all the way from the eye to the thalamus. The axon of the fourth neuron runs from the thalamus up to the visual cortex in the back of the brain, The first neuron, the rod or cone, is very small: it responds only to light shining directly upon it. Diffuse light is one easy way to stimulate a single rod or cone; if a small spot of light is used (such as when looking at a star), it must be centered directly upon the particular transducer cell. So the image of the visual world projected upside-down on the back of the eye is changed- into the voltages of a mosaic of transducer neurons. The second and third neurons in the chain start the process of comparing neighboring regions. A particular neuron responds best to a small spot of light, covering a number of adjacent rods and cones. If the light spot moves, the positive voltage changes into a negative one. If one moves the spot even farther away, the voltage becomes very small. So there is a doughnut-shaped map: the transducers in the center cause excitation, those in the surrounding ring cause inhibition.. What happens if both areas are illuminated, as happens with diffuse light? The positive and negative influences cancel - sometimes totally, as if the light were not there at all, sometimes only partially.
The fourth neuron, 4 in the thalamus, is even more discriminating against diffuse light. Essentially, the image of the visual world has been changed by emphasizing the borderlines between darker and lighter areas. Such a neuron may, however, respond equally well to a spot or line: it may not be able to tell the difference between such symbols as -, 1, /, -, +, etc. This is also true for the fifth neuron, located in the visual cortex. Starting with the sixth neuron, a voltage change can be evoked only if the orientation of a borderline is just right. If the animal is looking at a narrow line, such as the letter "1," a train of impulses may be evoked in some sixth-order cells. The line must be located in a certain part of the visual field, depending upon where the cell is located in the visual cortex, as there is a map of the visual world spread out across the visual cortex. Besides being in the correct place in space, the line must also be tilted at the correct angle. Orientation matters a lot to a sixth-order nerve cell (but any orientation will do in fifth-order cells and those earlier in the visual pathway). Some sixth-order neurons prefer vertical lines, others horizontal, others some particular angle. If the line is rotated 10 degrees from the preferred orientation, the impulses are likely to cease in that cell, but start up in another group of neurons.
The seventh-order cells often have even more complex properties. They, too, are particular about the orientation of a line but they are not as particular about its position in space. Some, called hypercomplex neurons, are particular about the line's length, however. Thus, some are specialists in short, horizontal lines. They could tell the difference between - and --. Thus the neurons of the visual system are not equally interested in everything one sees. They take apart the cameralike visual image presented to the mosaic of retinal transducer cells. It turns out that they take it apart in ways that extract information about borderlines between lighter and darker areas. If one looks at a checkerboard, most of the response of the visual brain is devoted to the lines forming the edges of the black and red squares. Many fewer cells seem to be needed to process the information about the centers of the squares. The visual cortex is wired up from birth to accomplish all of this; however, the wiring can be modified by the experiences of infancy, as the following case illustrates. Ross is six years old and cross-eyed. He seems almost blind in his right eye. But it now is too late to do much about it. Cross-eyed babies who see doctors regularly will be sent to an eye surgeon, as most doctors know that delays in correcting the defect will mean that the child may become functionally blind in one eye or the other. Lack of money in his family is not really the reason that Ross has now lost the sight of his right eye. Ignorance is expensive. His mother simply thought of correcting crossed eyes in the same cosmetic terms as fixing a crooked nose. That one shouldn't wait around too long with a cross-eyed child was just one of those empirical facts in medicine until a few years ago. Then, basic science finally advanced to a state in which techniques and knowledge were available to address the question: how does experience modify the brain's wiring? One of the first disorders which was illuminated by the research results was the mechanism by which the brain comes to ignore an otherwise good eye. 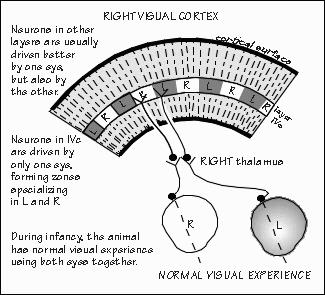
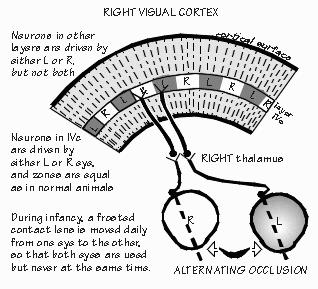
The two eyes see slightly different views of the world, as the reader can appreciate by alternately closing one eye and then the other, while reading this page. The "rangefinder" effect is quite useful for judging distances, at least out to 7 about five meters away (the length of an automobile). Not until the sixth-order nerve cell is the information from the left and right eyes combined, at least in primate visual cortex. At lower levels of the visual system, a cell either responds to light shown in the left eye, or to light shown in the right eye. In the cortex, there are cells responsive to both eyes. The visual cortex is wired up from birth with this ability, but it can be lost if the infant does not get a chance to use both eyes together. An infant monkey with a frosted contact lens, switched daily from one eye to the other, gets visual experience with each eye but the two eyes never get a chance to work together. The cells which would ordinarily respond to both eyes now seem to specialize in one eye or the other. Even if the contact lens is discarded and the monkey is then given normal visual experience for a long time with,both eyes together, the situation tends not to reverse: there will be very few coordinating nerve cells, permanently. There seems to be a critical period in infancy (in monkeys, about the first nine months after birth) during which experience with the environment can make permanent wiring changes, at least such obvious ones as these.
Such research, of course, has many implications for our own infants. Babies born cross-eyed have a problem similar to that of the infant monkey just noted; the two eyes never get a chance to work together. Rather than seeing double, humans with a squint will tend to suppress permanently the vision in one eye, becoming functionally blind in that eye (monkeys, interestingly, do not have this problem if made artificially cross-eyed; they can switch back and forth from one eye to the other). Such animal research has emphasized the need to correct cross-eyed conditions in infants as soon as practical; the relevant critical period in humans is probably the first two years after birth. There are also indications that there are critical periods for other brain systems besides the visual system. The timing of each system's critical periods is often different. Indeed, there is a hint that such critical periods may exist for human language. The existence of critical periods for normal brain development blurs the distinction between environment and heredity: now, early environment as well as heredity determines the brain machinery that is available for later interactions with the environment. As David Hubel noted, "Perhaps the most exciting possibility for the future is the extension of this type of work to other systems besides sensory. Experimental psychologists and psychiatrists both emphasize the importance of early experience on subsequent behavior patterns--could it be that deprivation of social contacts or the existence of other abnormal emotional situations early in life may lead to a deterioration or distortion of connections in some yet unexplored parts of the brain?" The current thinking among researchers is that during the critical period, there is competition among nerve cells in the visual cortex, a variation upon the survival of the fittest. If the frosted contact lens is left on one eye for a while during the first six months of life, the usual 50-50 representation of left and right eyes will change in the narrow band in layer IVc of the visual cortex where the fifth-order cells arc located. Fifth-order cells normally specialize in either left eye or right eye, never both. Normally, there is a territory about 0.4 mm wide containing only cells representing the left eye, then a 0.4 mm territory for the right eye, alternating left-right-left-right every 0.4 mm across the visual cortex. In the infant monkey with badly blurred vision in one eye from the frosted contact lens, the territories from the "bad" eye shrink and those for the good eye expand as if the active "exercised" cells had won a competition for space in the brain. 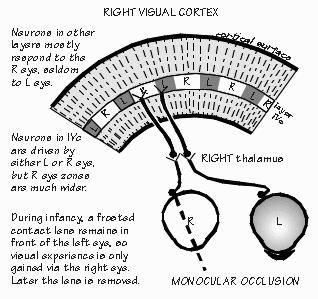
There is even a problem with animals growing up in vertically striped environments: some researchers report that .the orientation preference of the cells may shift, producing a higher than normal proportion of cells specializing in vertical orientations. As with other effects summarized above, there' is now a new round of scientific literature, pointing out the exceptions and variations and occasionally reporting failures to reproduce other researchers' results. (One famous failure to reproduce others' findings from cat visual cortex turned out to be due to slightly different breeds of Siamese cats in Massachusetts and Wisconsin; that discovery in turn has opened up new opportunities to study the wiring accidents during prenatal development from which these cats suffer). Thus a picture emerges of how the cells are wired together, how their internal electrical processing of information is carried out, where they send their new messages, and what happens next. |

 Continue to CHAPTER 12
Continue to CHAPTER 12
Notes and
References for this chapter Copyright ©1980 by |