William H. Calvin and George A. Ojemann, Inside the Brain: Mapping the Cortex, Exploring the Neuron (New American Library, 1980), chapter 5. See also http://WilliamCalvin.com/ Bk1/bk1ch6.htm. |
William H. Calvin
University of Washington |

|
6 Learning and Remembering: How Are Memories Recorded? |
©1980 by William H. Calvin and George A. Ojemann |
Studies of the psychology of memory in man and other animals indicate that memory includes several processes:
There are good reasons for believing that different mechanisms in the brain are used for short- and long-term memory storage. Short-term memory can be disrupted by something which interrupts brain function: a blow on the head, an epileptic seizure, or a brief interruption of the brain's blood supply. Immediately after recovery, the person may have trouble remembering what happened for the minutes (or hours, sometimes even days) prior to the brain disruption. This amnesia does not, however, extend to long-term memories. Even deep coma, which may silence much of the electrical activity of the brain, does not erase long-term memories. 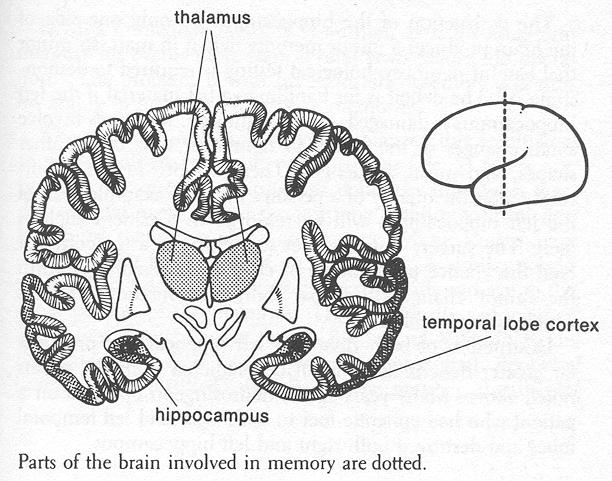
Long-term memories are not formed immediately after a new piece of information is acquired. New memories are "held" in short-term memory; animal experiments suggest that the more permanent long-term memory is formed gradually over a period of many hours or days. Disruption of brain function means not only that the memory is lost from the short-term memory mechanism but also that the permanent memory trace is imperfectly formed. Areas of brain involved with the short-term memory mechanisms have been partially identified. Regions of language cortex play a role, as noted in describing Neil's inability to recall a name following stimulation during the distraction. Portions of human thalamus seem to help focus attention, gating access into short-term memory and changing the "intensity" with which incoming information will be remembered. And many studies in animals and man have indicated that a structure on the inner side of the temporal lobe, the hippocampus, is centrally involved in the short-term memory process. The destruction of the hippocampus on only one side of the brain produces a minor memory deficit in man, so minor that careful neuropsychological testing is required to demonstrate it. The deficit is for handling verbal material if the left hippocampus is damaged; for the right side, the losses involve small changes in the ability to remember new faces, other shapes, and music sequences. These deficits do not significantly alter the quality of a person's life. For example, part of the left hippocampus will be removed in a patient such as Neil. The surgery thus involves something of a tradeoff. For Neil the chance to eliminate or control his seizures is worth the minor changes in verbal short-term memory abilities which will probably result. Destruction of both right and left hippocampus produces far greater deficits than might be predicted from the results noted above. Many years ago, a neurosurgeon operated on a patient who had epileptic foci in both right and left temporal lobes and destroyed both right and left hippocampus.
H.M.'s immediate memory is normal, as is his long-term memory for events more than several years before the operation. The problem is with his short-term memory and thus his ability to form new long-term memories. The neurosurgeon immediately publicized this result so as to prevent others from suffering a similar fate. H.M. is not, however, unique in suffering this deficit in short-term postdistractional memory and the resulting inability to form new long-term memories: there are patients whose strokes have destroyed the hippocampus on both sides. A similar condition, called Korsakoff's psychosis, occurs with damage to the inner portion of the thalamus. This is typically a disease of alcoholics, thought to be due to the lack of Vitamin B1 in their alcoholic diets (it has been proposed that alcoholic beverages be "fortified" with B1, in the manner of milk, to prevent the 12, 000 cases each year in the U.S. alone). These patients can often be discovered making up stories (confabulating) to cover up for their inability to remember. In contrast to these various brain areas which have been identified with the short-term memory process, it has been quite difficult to associate any particular brain structure with long-term memories. There seems to be no one area of brain where damage will leave a patient with the loss of long-term memory. The closest examples come from the patient with damage to the brain stem, who may go through a period in which he is awake and talking but cannot recall such simple long-term memories as his name and birthday. This, however, is apparently a failure of the retrieval mechanism, not the loss of stored information; the patient may remember everything later in recovery (except for being amnesic for the period just before and following the injury). Severe dementia, where many neurons in many cortical areas die, also produces a long-term memory deficit. Several decades ago, the Montreal neurosurgeon Wilder Penfield described some patients in whom electrical stimulation of the temporal lobe seemed to be able to "tap into" specific long-term memories. These observations were made during epilepsy operations such as Neil's.
Only a few patients reported such flashes of memory upon electrical stimulation; they often were those patients whose temporal-lobe seizures also evoked brief flashes of previous experience. One possibility is that the long history of seizures in these patients made some memories more accessible, so that even local electrical stimulation sufficed to evoke them. Some patients' memories were auditory (especially for left temporal lobe stimulation) and some were visual (especially for the right side). All had the quality that everything about a specific event was recalled, even down to the fine details and the emotional coloring. These findings led Penfield to propose that we have recorded all of our conscious experience (or at least all to which we were paying attention) somewhere in the brain, with the temporal lobe assisting in gaining access to that record. Just because we can no longer recall something does not .mean that it is lost, no longer recorded somewhere in the brain. The earliest studies on the psychology of memory were on people learning a foreign language. People who learn a language in early childhood, but who then switch to another language, often lose their ability to understand the early language. Yet when they go to relearn the language, they gain proficiency much more rapidly than others. This "savings" can be demonstrated for many kinds of learning. The degree of retention of a piece of information probably depends on a complex of things that occurred upon original acquisition: how intensely one attended to the material, and how much emotional response was stirred up. During the hours-to-days transition between short- and long-term storage processes, other factors are also likely to operate: how many associations did that piece of information make with other material already in memory, how much competition was there with other similar incoming information (the fewer things happening, the better might be the storage of what did occur), and the like. The number of times that the information is recalled and used affects the ease with which it may again be recalled. Finally, at the time of the recall attempt itself, how many associated items are already at the level of consciousness (perhaps in short-term memory again because of having been recalled minutes or hours previously)? Each of these processes probably affects the success of the recall attempt, each probably occurs using a different combination of brain areas, and each can probably be manipulated to improve long-term memory.
The foregoing discussion about memory indicates just how thin our understanding is concerning one of the most important aspects of the brain. There has been much speculation about memory mechanisms; what follows is a selection of the hypotheses and related facts. One possibility is that long-term memories may be coded in a string of molecules, in the manner of genetic memory, which uses DNA sequences. This would explain why longterm memories are so resistant to temporary disruptions of brain function; they are "built in" to neurons rather than depending on minute-to-minute electrical activity of neurons. But the DNA sequence is changed only by random mutation, not experience; experience might change RNA sequences or proteins. If this is the case, one might be able to extract the relevant chemical containing the information from the brain of a smart animal and give it to a dumb animal to make it smart. In the 1960s, there were reports of exactly this sort of result: worms trained to respond in a certain way to light flashes were ground up and fed to. untrained worms, who then learned the light response faster than expected.' This led to the obvious suggestion that students would no longer need to attend lectures if they ate the professor. There are other difficulties with this theory, too. While protein synthesis in cerebral cortex may be necessary if a long-term memory trace is to be established (applying inhibitors of protein synthesis after exposure to new information interferes with its retention), there are many technical issues associated with such experiments which complicate their interpretation. Besides changes at the biochemical level, it is thought that anatomical alterations might occur in neurons following learning. The anatomical change is unlikely to involve just one neuron or synapse: with aging, neurons are lost but memories are not similarly lost in a piecemeal fashion. A given neuron probably cooperates with many other neurons in forming the trace of a particular memory. As with people trying to push a car out of a ditch, no one person may be essential; there may be a minimum number of people needed, and it is likely that the task will be accomplished more efficiently if everyone works cooperatively. A given memory may involve changes in many neurons, perhaps involving a number of different specialized regions of the brain. 
Our thinking about possible memory mechanisms is strongly influenced by the analogies which science and technology have provided us thus far: the term "laying down the memory trace" probably originated by analogy with a groove being cut on a phonograph disk. Then came the digital computer having both short- and long-term memory mechanisms: when the power fails, the information in the transistorized active registers is lost, but that in the magnetic core survives. The analogy is defective in that computers store a given piece of information in a discrete pigeonhole. The hologram has provided us with a better, though imperfect, analogy for the distributed aspects of long-term memory storage. Evolutionary principles, where random genetic variations are selectively conserved to form new animal species, also provide us with an analogy for the formation of new long-term memories by conserving some random connections. While the number of neurons does not increase after birth, the size of the individual neuron changes greatly. In infancy, some neurons send out many short branches (called spines) for a while; this is followed by a period in late infancy when the number of spines declines by half, as if they had been pruned (see p. 126). Another model for long-term memory storage in the adult brain has been based upon similar pruning. It assumes that even adult neurons are continually budding and sending new branches out to make random connections with axons in the vicinity. Men some new event happens and the brain processes the information, many neurons are activated. Suppose that the new synapse happens to participate in this process. Imagine a pruning process that breaks new connections which were not utilized in the preceding day. This would leave intact those synapses which were used. The anatomical substrate for the memory would therefore take some time to be "developed," as in the photographic development of a negative where unexposed silver grains are removed. Thus, laying down the permanent memory trace might simply be a matter of conserving those randomly made connections which happen to participate in responding to a new event. Short-term memory might involve the temporary strengthening of a synapse (see facilitation in Chapter 10) following extensive use by increasing for a day the amount of neurotransmitter released by an impulse. Indeed, the facilitation changes in the synapse might be what protects the new synapse from the pruning process. Such a theoretical model helps to explain why, although short-term memories are easily disrupted by massive activation as in seizures (analogous to fogging the film), long-term memories could require some time to develop and be disruption-proof thereafter.
|


 Continue to CHAPTER 7
Continue to CHAPTER 7
Notes and
References for this chapter Copyright ©1980 by |