William H. Calvin and George A. Ojemann, Inside the Brain: Mapping the Cortex, Exploring the Neuron (New American Library, 1980), chapter 9. See also http://WilliamCalvin.com/ Bk1/bk1ch9.htm. |
William H. Calvin
University of Washington |
|
9 Paying Attention: |
©1980 by William H. Calvin and George A. Ojemann |
The reason that the activating system of the brain is so effective at spreading seizures around is its widespread connections to most parts of the brain. It has two components, arousal and selective attention; together, they are essential for higher brain functions. Different sections of the arousal component of the reticular activating system regulate the complex processes of sleep. These sections are all located in the upper part of the brain stem. One section triggers deep sleep, also called "slow-wave sleep" because of the changes in the brain waves that go along with it. Normal awake resting brain waves have many oscillations at about eight to twelve cycles per second, called the "alpha rhythm" (a favorite of those who like to use biofeedback to condition brain waves). When the reticular activating system arouses a person, the alpha rhythm is replaced by activity that oscillates much faster and over a smaller voltage range. During deep sleep, the waves are much bigger and slower, oscillating one to two times a second, and leading to the name "slow-wave sleep." This deep sleep is periodically interrupted by rapid-eye-movement (REM) sleep. These REM periods are triggered by another section of the reticular activating system. During REM, the brain waves oscillate more quickly, as though the person were awake; the eyes move about rapidly beneath the closed eyelids (hence the name), although other muscles are relaxed. If the person wakes during REM sleep, he often reports dreaming. REM sleep seems to be essential: if a subject is awakened every time he starts a period of REM sleep, there will be more REM sleep the next night. If such REM deprivation is kept up night after night, the subject's daytime performance will deteriorate much more than if the awakening had occurred in deep sleep. But why we need REM sleep, with its dreaming, is unknown. The arousal part of the activating system is located in the core of the upper portion of the brain stem. This site is rather vulnerable to damage from head injuries. This is probably because the brain stem acts much like the stem of a mushroom. During a blow to the head, the cerebral hemispheres may oscillate back and forth and put stress upon the brain stem, the way that a mushroom stem would suffer if the top of the mushroom were kicked. Brain-stem injuries often lead to the loss of consciousness because of trauma to the activating system; while the damage is often reversible, permanent coma sometimes results. The second component of the activating system involves a more refined process: selective attention, rather than generalized arousal. This selective attention allows the discrimination of meaningful from unimportant stimuli: a mother responds to her baby's cry but not the noises of other children playing. It allows sustained concentration on one task, such as reading this chapter, with only a novel or imperative stimulus able to distract, such as a sonic boom or a call to dinner. Part of this component of the brain's activating system is located in the thalamus. It is the system altered by electrical stimulation during thalamic operations for Parkinson's disease, stimulation that shifts attention from the patient's internal world to what is coming in from the external world. This attentional shift enhances memory for what is coming in, while making it harder to call up information already in memory. The left thalamus in man has this property for verbal material, the right for visual-spatial information. In animals, this selective attention system shows little evidence of lateralized function. It is tempting to speculate that this system should play a major role in conscious experience. It should determine to what we attend in the environment and the intensity with which it is remembered. When it switches away from external to; internal stimuli, associations already in memory become Available to compare with external stimuli, and as a source for internal modeling of different response strategies before taking action. We often rehearse such internal strategies: how to respond to the boss when told of a promotion; what is the best way to get from the freeway to the restaurant for tonight's dinner engagement; how to design an experiment to determine the role of human alerting mechanisms. Each of these internally rehearsed models generates new associations from short- and long-term memory. This internal rehearsal often occurs while, elsewhere in the brain, external stimuli are receiving appropriate responses: the to-be-promoted-secretary types a letter, the driver continues down the freeway avoiding other cars, the experiment designer finishes showering. The selective attention mechanism seems to be the system turning different brain areas on and off to the external or internal world. The internal-world aspect of conscious experience is enormously resistant to experimental study. Each of us knows that we have such an inner world, complete with "inner speech" and private ideas, but how does one establish the nature and existence of that inner world in someone else, or in another animal species? One can imagine the diseases that would result if this selective attention system malfunctioned. With a minor disturbance of the system, the ability to hold attention on the external stimulus at hand would weaken, so that attention could be easily diverted by other external or internal stimuli. Such a defect is commonly seen in children with learning difficulties, children often labeled as "minimally brain damaged" or "minimally learning-disabled." Indeed, a proportion of these children show other neurologic signs suggesting that the thalamus or striatum may not be functioning normally. Patients with hemorrhages in the left thalamus show a somewhat different defect. Attention can be sustained upon the external world, but things (words) from the internal world seem to intrude into external speech in an uncontrolled way, much to the patient's surprise and frustration, as in our patient who kept having the phrase "affirmative action" intrude on his attempts to name objects. If the failure of the selective attention system were complete, one would expect a loss of all attention to external stimuli; the brain would be continuously turned in to its internal world. That seems to describe what often happens in autistic children, who show minimal or no attention to external stimuli. They act as though they are continuously attending to something inside themselves. But anatomical studies of the brains of autistic children show no anomalies, not even in striatum and thalamus, the areas where the selective attention system is thought to be located. Perhaps these studies have not looked for the right things, or perhaps our intuitive idea of what -a damaged selective attention system would do is wrong. A partial failure of the selective attention system might result in something in between minimal brain damage and autism. This seems similar to one of the defects described in schizophrenia. These patients "are brought to a topic totally irrelevant to the subject at hand, by any accidental thing that happens to affect their senses. . . . The normal directives through questions from without and purposive questions from within are incapable of holding the train of thought in the "proper channels." Schizophrenics may also blur the distinction between internal and external worlds. This is in contrast to the patient with a thalamic hemorrhage who seems to know this distinction clearly, even though the internal world intrudes on the external. Indeed, schizophrenia is a many-faceted disease, perhaps with multiple causes involving several different brain areas in different cases. Defects in selective attention are, at best, only part of the problem. A situation similar to acute schizophrenia can be produced in a normal person by an overdose of the drug amphetamine. Study of similar amphetamine overdosage experimental animals has uncovered a clue to a neurotransmitter abnormality in the parts of the brain concerned with selective attention, a clue that eventually may be important in understanding what is wrong in schizophrenia. The culprit again seems to be the neurotransmitter dopamine-not too little dopamine, as was the case in Parkinson's disease, but rather too much effect from a normal amount.' A brief look at the connection between nerve cells, the synapse, and the ways it can be fooled illustrates how this seems to come about.
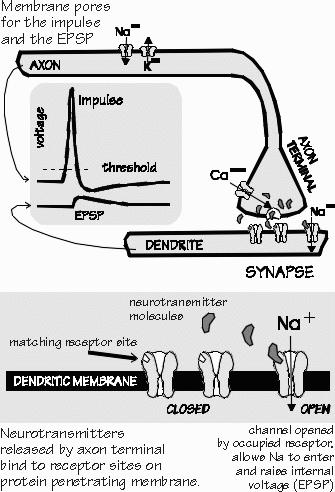
There are a number of steps in the process which one neuron uses to talk with another, a process called synaptic transmission. An electrical impulse in the first neuron travels down to the end of the axon, where there are regions specialized for releasing neurotransmitter molecules from the cell. A certain number of molecules are released by each impulse; the release process is triggered by calcium entering the cell following the voltage change of the impulse. The molecules start diffusing away from the cell membrane after their release, but the next neuron is just a short distance away, across what is called -the synaptic cleft. The surface of the next neuron at the synapse is quite specialized for detecting the neurotransmitter molecules. There are a number of sites called receptors, where a chemical bond can be formed between the neurotransmitter molecule and the receptor molecule. The combination molecule then unlocks the membrane of the second neuron to allow certain substances through, such as sodium, potassium, or chloride ions. These ions, in turn, change the voltage of the second neuron. So the first cell releases "keys," the second cell has a number of "keyholes," and a key which fits the lock opens a "gate." There are also mechanisms in the synaptic cleft for cleaning up neurotransmitter molecules. Rather like a vacuum cleaner for keys, these "Inactivation" mechanisms prevent some of the transmitter molecules from ever reaching the keyholes I'll the first place-indeed, they "suck the keys" back out of the "keyholes" after a short time, which results in the gates locking again to stop admitting chemicals to the second neuron. This is one way in which the effect of the impulse is limited in time: an impulse may last one millisecond itself, but its effect upon the second neuron via the neurotransmitter it releases may either be brief or long-lasting. At one time, it was thought that in schizophrenia, neurons released too many dopamine molecules each time an impulse arrived at a synapse. However, the supporting evidence for that idea has been mixed. Autopsy study of the brains of some schizophrenics suggests that there are many more receptors for dopamine in the striatum than in the same brain area of normal people. There thus may be an excess of "keyholes," rather than more "keys." And since the usual situation is to have more neurotransmitter available than receptors (more keys than keyholes), the presence of more receptors means that each impulse has a greater effect on the next neuron. In other diseases, too little neurotransmitter is produced: there are too few "keys," as in the shortage of dopamine in Parkinson's disease. In still other diseases, there may be too few keyholes. Apparently this happens in myasthenia gravis, in which the body's immune mechanism runs amok and destroys 80 percent of the receptor sites ' at the nerve-muscle synapse. There are problerns when the cleanup process fails, so that neurotransmitters stay around too long. Some poisons work this way, such as nerve gases and the pesticides. Other poisons such as curare act as if they were keys which fit the lock but don't turn, their presence in the keyhole keeping the right keys from entering the lock.' Many drugs more gently affect the synapse by changing the number of keys, by partially occupying keyholes, or by slowing the cleanup. In one way or another, the effect of the impulse is being modulated to alter the strength of the functional connection between the two neurons. From such synaptic alterations come the hallucinations of LSD and the therapeutic actions of levodopa. What drugs modify the synapses in the memory or selective-attention pathways? No one knows. |
 Continue to CHAPTER 10
Continue to CHAPTER 10
Notes and
References for this chapter Copyright ©1980 by |