[How] we come to analyze the world, without postulating the presence of a non-material central agent or homunculus... is a problem that is rarely faced by
neuroscientists.... Perhaps the reason for this serious failure is that
neuroscientists mostly think about individual units but not about the
population
of neurons and their connections, the synapses of the brain.
the English neurobiologist J. Z. Young, 1988
To learn is to eliminate.
the French neurobiologist Jean-Pierre Changeux, 1983
Surely there is something better than an infinite regress of little people inside the head. Or the
infinitely regressing agendas of "reductionism forever," constantly changing so that one never
answers the interesting questions about how the mind works.
Well, one thing that's better is the bookstore. The Shining Sea Bikeway is the only path I know
that has excellent bookstores at each end, both in Falmouth and in Woods Hole. No matter
which end you start from, you can take a break in the middle of your round trip by sitting down
and browsing through the new books. A truly enlightened trail, with some foresightful soul
having provided park benches every mile and, not far from each end, a truly civilized bookstore
with a readers' table and chairs. And J. Z. Young's new book, Philosophy and the Brain.
"Jay Zed," after providing the membrane biophysicists with the squid giant axon (he and Keffer
Hartline did the first recordings from it here; Hartline was surprised, never having seen such a
large electrical signal before, and suspected faulty equipment initially), went on to study the other
familiar cephalopod mollusk, the octopus. It is the most intelligent of the invertebrates,
sometimes compared to a laboratory rat in its behavioral versatility. And after modeling the
octopus's visual memory, he suggested that memory may work, in part, by eliminating some
neural connections -- by carving away material, in the same way those carved figureheads for old
sailing ships were created.
This was somewhat heretical: Everybody was used to thinking of growth and memory as a
process of adding on material, the way children with modeling clay are likely to make a figurine
by padding it out. Neurobiologists are like everyone else, always filling up one file cabinet after
another, and when we said that memory was a cumulative process, we usually meant it in both
senses of the word. J.Z.'s proposal had the interesting property of suggesting some limit to how
much information could be stored with such a process: After all, a carver who doesn't know
when to stop soon runs out of wood. If you lived long enough, you might run out of brain! And
surely consciousness before that. You can see why the idea didn't immediately catch on.
Because new memories had to be somehow superimposed on old memories, one imagined
smaller figures being carved into larger ones, as in graffiti on a wooden figurehead; from a
distance, it would loom like one thing, but focusing up closer would tell a different story. So
adding on new memories had to conform in some way to the old memories: after all, carving a
whale does make it hard to superimpose the form of a giraffe on it. Reorganizing your
memories, in the manner that I periodically reorganize my file cabinets, might be a little hard if
the information was stored by carving. Certainly neural carving raised the possibility that some
experiences could produce irreversible effects.
Much to my surprise, carving connections has provided one of the key insights required to
understand how consciousness committees function, and for why one doesn't need labeled lines
to get Grandmother's face recognition. It's all because of the Mexican hat -- what was originally
called lateral inhibition by its neurophysiological discoverers, Georg von Békésy, Keffer
Hartline, Floyd Ratliff, Stephen Kuffler, Robert Barlow, and such.
To begin personally, on a confessional note, I was at one time, at my onset, a
single cell.... I do not remember this, but I know that I began dividing. I have
probably never worked so hard, and never again with such skill and certainty....
At one stage I possessed an excellent kidney, good enough for any higher fish;
then I thought better and destroyed it all at once, installing in its place a neater
pair for living on land. I didn't plan on this when it was going on, but my cells,
with a better memory, did.
Thinking back, I count myself lucky that I was not in charge at the time. If it
had been left to me to do the mapping of my cells I would have got it wrong,
dropped something, forgotten where to assemble my neural crest, confused it. Or
I might have been stopped in my tracks, panicked by the massive deaths, billions
of my embryonic cells being killed off systematically to make room for their more senior successors, death on a
scale so vast that I can't think of it without wincing. By the time I was born,
more of me had died than had survived. It is no wonder I can't remember;
during that time I went through brain after brain for nine months, finally
contriving the one model that could be human, equipped for language.
the American physician and essayist Lewis Thomas, 1987
IRREVERSIBILITY has been an important idea about human beings for a long time. The Jesuits
thought that they could produce lifetime obedience with suitable indoctrination of little boys;
Charles Darwin noted that "a belief constantly inculcated during the early years of life, while the
brain is impressible, appears to acquire almost the nature of an instinct." Defense attorneys
invoke such reasoning to attempt to excuse their clients' actions, saying that their behavior was
all the manufacturer's fault, i.e., the school system and "society" in general. Some psychologists
claim that the child's brain is infinitely malleable by the environment in which it grows up -- but
usually recognize that adults are more likely to become set in their ways as they grow older.
Whatever the extent of the plasticity in various periods, it certainly seems likely that something
so important will be carefully regulated, probably on a day-to-day basis (or night-to-night -- one
proposed function of sleep is to adjust what is passed from temporary to long-lasting memory).
Behavioral plasticity may be restricted to some "critical periods" in development; in certain
respects, it really matters what your experiences are during some years. If the two eyes don't get
a chance to work together within the first two years of life, chances are that one eye will become
functionally blind; there will be nothing wrong with it optically, but it will act as if disconnected
from the higher centers of the brain (this is the reason why cross-eyed babies need such early
surgery).
Something as well learned as a schema becomes the modal schematic of a series of similar
experiences: We no longer remember when we first encountered the letter A, or when we took
our first step. A snapshot schema, however, is a record of a unique event (in my generation:
what you were doing when you heard about President Kennedy being shot). Unless there is some
way of freezing it (the way those infantile memories of binocular experience were frozen by
myelination and synaptic editing), we might expect such memories to be malleable. We shape up
schemas to be the average shape of a lighthouse or the defining character of a Picasso. The lack
of "fixing" means that every time we reactivate the schema (as when we recall it "to mind" in the
absence of the original), we stand some chance of modifying the memory.
History is what you remember, and if you don't think it's being revised all the
time, you haven't paid enough attention to your own memory. When you
remember something, you don't remember the thing itself -- you just remember
the last time you remembered it.
the Grateful Dead songwriter John Perry Barlow, 1984
MALLEABILITY IS THE FLIP SIDE of irreversibility: Our memories are sometimes more
modifiable than we imagine. Indeed, each time we recall something, we have an opportunity to
modify that memory. This means that we can readily fool ourselves, unless we have disciplined
ourselves to keep fantasy separate from reality. Even then, "brainwashing" techniques (of which
some religious-conversion practices are the most familiar) may persuade us that the exact
opposite is true. Ordinarily, human memories are pretty good, and we have come to insist (as a
matter of social policy) upon one's responsibility for telling the truth -- but there are no
guarantees biologically, just as there are no guarantees against mental illness.
So how does this malleability, this plasticity, of memory arise -- and how is it occasionally
"fixed" to resist further change? There are no general answers yet, but a spectrum of phenomena
has been uncovered, ranging from dramatic examples of carving during childhood to subtle
modeling of interconnections during adult experience "permitted" by neuromodulators.
I was reminded of all this when I sat in on the neural systems course and heard Patricia Goldman-Rakic lecture on the frontal lobes (yes, indeed; frontal lobes have finally made it to MBL!). We
usually think of the process of growing up as one of growth, of adding on to the body and filling
it out with additional material. But development is also a process of removing some material.
This isn't the familiar old saw about "you lose ten-thousand nerve cells every day" (the experts
now say that isn't particularly true in higher centers, that nearly all the cell death in cerebral
cortex is accomplished before you are born, even though some subcortical structures such as the
substantia nigra lose a lot of cells during life -- and excess loss is a component of what we know
as Parkinson's disease). But there is an evolutionary version of the carving principle, some
neurobiologists now saying that new neural structures deep in the brain have been created by a
process of removing intervening cells to define their shapes better, that this is how differentiation
of "subdepartments" occurs.
Nerve cells also die during one's lifetime, of course, as when injured by a bruise or the loss of
oxygen supply or by escaped hemoglobin (when blood cells rupture, and the arterial walls as
well, the hemoglobin may come into direct contact with nerve cells and kill them). These local
areas of damage may never be noticed, as adjacent areas take over their function so well. When
large areas of brain start losing a lot of nerve cells, however, you start having problems with
memory, reasoning, speech, and all the rest.
There was an old Gary Trudeau cartoon posted on the bulletin board in the student lab for the
neural-systems course, from the series "In Search of Reagan's Mind," where the investigative
reporter is wandering around inside the convolutions of a seemingly empty frontal lobe, shining
his flashlight here and there and exclaiming, "Where is Ronald Reagan?" But neurologists don't
find that so funny: Every day they look at magnetic resonance scans (those computerized
pictures that seem to slice up the brain; they're particularly useful for judging the size of various
structures) from patients who are losing a lot of frontal lobe and getting senile, exhibiting many
of the signs and symptoms that Elaine had for a month and then got over.
Alas, the senile patients with such massive loss don't recover: once brain cells are gone, they're
not typically replaced (unlike the blood cells, which are totally replaced every 120 days, or the
intestinal lining which is replaced every 3 days). Neurologists look at the brain images and see
the infolded cortex developing wide, unoccupied valleys, and they sigh regretfully. In a normal
brain, those valley walls are pushed together so tightly that you cannot see down into the groove.
Strangely, many magazine photographs of "normal brains" are really the brains of people who
had senile dementia -- that's because art directors like exaggerated features that reveal the deep
folds, and so they pick the pictures that "look best." But those brains had been carved from
within by the disease process that destroyed most of the cortical nerve cells and shrank the brain.
You wouldn't want to have a brain like the ones that they, in their ignorance, like to picture.
Is the disease process an exaggeration of normal developmental processes, something like
cancer? Cancer is wild proliferation of cells, adding on more cells uncontrollably to create
tumors and infiltrate other tissues, the way that strip cities invade farmlands insidiously. Is this
loss of cells in senile dementia a disorder of a later stage of development where editing, not
addition, is the dominant feature? That is just one of the many reasons why so many people
around MBL are studying developmental processes in biology, trying to learn the normal rules of
the game so as to better understand how things go wrong.
When one sees a process in biology, such as cell proliferation, one is almost sure to see one or
more additional processes, such as cell editing, that oppose it. Everything is usually a tug-of-war,
with net movement occurring only when strengths no longer balance. Yet the two opposing
processes are seldom symmetrical like the tug-of-war with a dozen people on each end of the
rope: In biology you find situations more like a winch being used on one end of the rope, people
on the other. Push and pull often come about in different ways, and it may not be obvious what
is being "balanced."
We learn... that there's a utility in death because... the world goes on changing
and we can't keep up with it. If I have any disciples, you can say this of every
one of them, they think for themselves.
the pioneer neuroscientist Warren S. McCulloch
EDITING CONNECTIONS during childhood is a much less destructive process than the loss of
entire cells during prenatal development or senile dementia. What we are talking about here is
not nerve cells dying, but selectively breaking half the interconnections between cortical nerve
cells.
As we grow up, we lose close to half of all the interconnections in our cerebral cortex -- we
gained connections until about eight months after birth, but after that comes this net loss. It can
be even worse for some long-distance connections, e.g., the connections between the monkey's
left brain and right brain decline by 70 percent between birth and sexual maturity. Mammalian
brains have connections from all areas of cerebral cortex to the spinal cord at birth but, by
maturity, all have been withdrawn excerpt for those from the usual somatosensory, motor, and
premotor cortical areas.
That is a lot to lose; if you'd told me (or any other neurophysiologist) this fact maybe two decades
ago, I wouldn't have believed it. Nerve cells seem to start out by making lots of connections with
other nerve cells (not quite "everything is connected to everything," but much more widely than
anyone thought) -- and then something edits them, disconnects quite a few, shaping up the child's
mind by whittling away. There's an important principle here: Make lots of overlapping
connections, then narrow them down somewhat -- but not all the way down to unique "labeled
lines." The necessity for such a disconnection principle was recognized a quarter-century ago by
a philosopher, Daniel C. Dennett, in his 1965 doctoral dissertation at Oxford; he even recognized
the analogy to biology's pre-Darwin convergent selection:
What is needed is for some intra-cerebral function to take over the evolutionary role
played by the exigencies of nature in species evolution; i.e., some force to
extinguish the inappropriate.... This would have the effect of pruning the initially
unstructured connections along lines at least compatible with and occasionally
contributory to the appropriate inherited links already endowed by species
evolution....
The process is a repeated self-purification of function, gaining in effectiveness as
more and more not inappropriate structure becomes established.
Least destructive of all would be simple modifications in the strength of the synaptic connections
between cells, being able to diminish the strength to nothing without actually disconnecting it (a
"silent synapse"). Maybe physical disconnection is one way of "fixing" the memories encoded
by such reductions-to-nothing; if the connection remains physically there, some future retuning
of the system might destroy the old memory that relied on the weak connection.
So there seem to be a variety of ways of editing brains: killing whole cells (as happens in
prenatal development and senile dementia, and possibly in songbirds that learn a new song every
year), disconnecting some interconnections between selected cells (as happens during the tuning-up to the environment seen in childhood), and simple increases and decreases in synaptic
strengths (as certainly happens in short-term memory throughout childhood and adult life).
Long-term memories, of the multitrial varieties we call schemas, likely involve both altered
synaptic strengths -- and sometimes the creation of additional synapses by an existing cell
budding off a new axon branch and attaching to another cell.
New synapses? All that the childhood halving of synaptic numbers in cerebral cortex means is
that there is a difference between the rate at which new connections are being made and the rate
at which old synapses are being disconnected. Up to eight months after birth in humans, the
creation rate exceeds the destruction rate; afterward, slightly more are disconnected than new
ones are formed. But no one knows how many synapses are being destroyed in the average
week: All we know is the cumulative difference between creation and destruction rates, which
yields a 35-50 percent loss during childhood. We have no way of tracking individual cortical
synapses over time in a given animal, though brain imaging techniques that visualize proteins
involved in making new synapses should eventually give us a clue about creation rates.
Sprouting to make brand-new connections gives us an additional process to modify for memory's
sake: We have little idea of how frequently this happens in adult life, or if there are favored sites
for sprouting, or how it might be regulated.
Worrying about fixing snapshot schema may, of course, be needless: There is no evidence that
humans were designed by evolution to be faithful recorders of events. It is true that one-trial
learning exists, particularly for the tastes of foods that make you sick. But there is nothing about
that which says that the memory must remain forever fixed. Modification is probably the rule,
not the exception.
Nothing seems more possible to me than that people some day will come to the
definite opinion that there is no copy in the...nervous system which corresponds
to a particular thought, or a particular idea, or memory.
the English philosopher Ludwig Wittgenstein (1889-1951)
Information is not stored anywhere in particular. Rather, it is stored everywhere.
Information is better thought of as "evoked" than "found."
the American cognitive scientists David Rumelhart and Donald Norman,
1981
POINT-TO-POINT REPRESENTATION is the notion that maps are connected in orderly ways,
as by a pipeline from the tip of your little finger to your somatosensory cortex's little finger
region. Or from a photoreceptor on the retina to the corresponding place in the visual cortex's
map of the visual world that "represents" that direction from the eye. I suppose that it was
reasonable to expect this, but we've known for a long time that it wasn't that simple. For
example, the visual world is represented by several hundred million photoreceptors in the two
eyes, but they have to get funneled down into several million optic-nerve axons. So, we said,
maybe the fine grain is only several million points instead.
But then it turned out that we can detect line spacings that are finer than the spacing between
photoreceptors (what is called "hyperacuity"): We're even better than hundreds of millions!
How can this be? It is because "Mexican hat" committees do the job, not pipelines. It is a
population of cells at work, not just a single cell lighting up while the others keep quiet.
The only way to understand how information is stored in the brain may be to understand what the
information is being used for; that's always been obvious for learning new skills but isn't so clear
for our more detached kinds of knowledge such as words. Certainly for the more familiar kinds
of computers, you need not understand the program to understand storage techniques -- but
processing and storage are all mixed up in nervous systems. The brain circuitry that analyzes the
information is likely to be used to store it as well. So all that pruning of synapses is likely to
subserve an analysis or performance function as well as a storage function. If many-to-many is
the circuitry connecting sensory and performance regions of the nervous system, rather than
many-to-one-to-many, we will simply have to learn to think in population terms -- just as Darwin
did when contemplating transformations of one animal species into another.
The most familiar transformations in our everyday experience are associated with hearing: The
treble and bass controls on a radio that augment or reduce the high and low ends of the spectrum.
Some hi-fi setups even have equalizers so that a half-dozen different parts of the spectrum can be
adjusted separately. You transform what's really there into an altered version that is more
pleasing. The brain is doing such transformations, and adjusting them, all the time -- but
internally, without twiddling knobs.
Our nervous system is indeed in the business of transforming things, not in the TV camera's
"faithful reproduction" business. Sometimes what you see doesn't correspond perfectly to what
you feel touching the same objects (so which is "reality"?). What we "see" when we look at a
seashore scene is not what a TV camera would record -- it is subtly different because of the
transformations taking place that help extract the information our brains need to make decisions.
Some of the differences from reality are simply called "illusions," as they are unwanted side
effects of the processing: Look at a waterfall for a moment, at the waves of water tumbling
down, and then look at the trees nearby. The trees will seem to be moving upward! Look
between your fingers at a bright light: You will see some little black lines partway between your
fingers. They are not interference fringes but illusions called Mach bands, a side effect of a
contrast enhancement transformation that occurs at several levels of visual processing in eye and
brain. When we say we "see" something, we are simply reporting on one intermediate stage of a
multi-stage set of transformations -- probably just the stage that is accessible to our language
cortex.
The transformations aren't always the same: they are adjusted as conditions alter. In the
moonlight, one can see pretty well (though in monochrome). But try to read a newspaper in such
light and you'll discover that the type is indistinct, as if irredeemably out of focus. Try to catch a
thrown ball at dusk, and you'll discover that your visual images are too slow to keep track of ball
position. The retina has readjusted some of the inhibitory mechanisms in the retina that enhance
both spatial and temporal resolution, choosing to improve low-light sensitivity at their expense.
If you were missing those spatial and temporal transformations in the daylight, you wouldn't be
able to either to read or catch balls: What we normally "see" is enhanced in some respects,
degraded in others, and has unrealistic features added. So much for "reality"!
And, of course, different animals are tuned up in different ways. Those primitive Limulus
wandering around offshore have about ten eyes, positioned at various strategic points around that
horseshoe-shaped shell (including one on the tail that specializes in day-night rhythms!); most of
the eyes probably use inhibition for contrast enhancement. And Limulus is extraordinarily good
at detecting faint shadows even in moonlight, even when two stories deep offshore Stony Beach.
Robert Barlow, a second-generation neurobiologist who studies lateral inhibition at MBL (in
neurobiology, one can trace genealogies back to just a few pre-1940 workers: Barlow was a
student of Keffer Hartline's, whereas my wife and I are third-generation neurobiologists, both
students of another student of Hartline's, Charles F. Stevens), says he can swim around in scuba
gear at the full moon, and when his dim shadow falls across a Limulus on the dark bottom, the
animal will change course. He can make it crawl along a zigzag course by simply casting a
shadow on its left side, then its right side, etc
I JUST SAW A FISHING BOAT return with a whole class of MBL students packed into its
stern, standing room only. I can't believe they collected anything, as they were only out one hour.
And there wasn't room for any fishing gear on the stern, so tightly packed was it with students.
Sightseeing? Well, at least they saw the salt water from a boat -- that's more marine experience
than most students get here these days.
MBL is an oddity among marine stations: The staple course taught by nearly all marine labs is
comparative invertebrate zoology. But it's not taught at MBL anymore. At other marine research
stations, the tide tables are prominently posted and the researchers are likely to go out and collect
their own animals from one or another of a fleet of small boats tied up at the dock. Life at such
labs revolves around the tides. One sees rubber boots, set out to dry alongside the special nets
and traps, diver's tanks and weighted belts lying out on the docks, instructions posted about how
to reach the nearest decompression facility in case struck by the "bends." But the Marine
Biological Laboratory is rather urban: The animals appear in aquarium tanks, delivered by ex-fisherman employees. With the exception of some people such as Bob Barlow, if you asked the
typical MBL researcher when the best low tide of the season was going to be, you'd draw a blank
stare. Or maybe, "What tides?" The MBL is now "marine" only at one remove for many of the
researchers.
Some of the research at MBL could now be done in the middle of New York City with the
animals delivered by air freight in Styrofoam picnic chests -- an option that was not available in
MBL's formative years. There are still many notable exceptions to that statement, such as the
biophysics done on the fragile squid and the developmental biology on various eggs of marine
organisms, and that research remains the hard core of MBL biology. But there are certainly some
researchers around here who could get by on ice chests and couriers.
So why do they continue coming to Woods Hole? MBL is an expensive item for most
researchers, not only for their supporting budgets but personally, as the housing around here is
(thanks to the better-paid Boston bankers competing for it) so expensive that researchers' savings
accounts suffer. It can't be scenery -- going to the beach is far easier and cheaper elsewhere on
the East Coast, and the people who come here to work typically go elsewhere for serious
vacations. So what is the real reason that so many researchers still go to all the trouble of crating
up their labs, suffering with the rental truck, the sore back, and the equipment that inexplicably
stops working when disturbed -- and then repeating the tasks several months later?
It's that MBL is, scientifically, a very special place quite aside from the setting and the animal
availability that were associated with its origins. You learn important things here in three weeks,
things that you'd seldom learn back home or at a convention center meeting amid ten thousand
milling scientists. People are set up and working here; you can get a demonstration of something
ten minutes after you hear about it, check things out in the extensive library after lunch, and try
the modification out on your lab rig that afternoon. The rumor mill in techniques and
preliminary results is a fundamental part of doing science, even if it is hard to document. Its
buildings and animals are vital, but it is as a social institution that MBL is so influential in
biological science. No other marine lab has a comparable level of free exchange of important
ideas, certainly not built into its basic program of courses, conferences, and rich diet of special
lectures.
One of the pleasures of summer in Woods Hole is the breadth of evening lectures not only on
science but on art, history, and public affairs. There are concerts several times a week (the
philosopher Geoffrey Hellman just played the Brahms' Intermezzo in B minor last night as the
encore to a superb piano concert of Mozart, Berg, and Beethoven). "Try to learn something
about everything, and everything about something" was Thomas Henry Huxley's epitaph; he
would have liked Woods Hole.
What Woods Hole has now is, I suspect, partly a legacy of generations of nonworking wives with
a lot of time and energy to organize. This was a form of scaffolding, now largely removed as
most educated women pursue their own careers, but historians may come to see educated
nonworking wives (and the occasional nonscientist husband of an MBL researcher) as essential
intermediaries for the present cultural milieu of Woods Hole, what makes the social life here
more than a place to meet people, more than the usual buffer to diffuse fatigue and hostilities.
They've created a milieu that makes learning something about everything extend well beyond its
usual liberal arts boundaries, and often bridge C. P. Snow's two cultures with grace. The number
of families around here with second- and third-generation scientist-physician-musicians is one
indicator of its success.
Buildings are buildings, but this feat of social engineering was far harder to achieve. And it is
potentially fragile, capable of being wiped out by empire-building government agencies who
want to keep "their people" on a short leash tied to Washington, D.C., or by an economy-minded
Congress bent on cutting costs by consolidating facilities for "efficiency." MBL has no
institutional backing -- it's an independent nonprofit corporation owned by its seven-hundred
scientist members, and its finances are always worrisome. But places like MBL are not defense
contractors producing products, nor even designers with a definable output in terms of blueprints
-- they are think tanks, first and foremost. In the physical sciences, think tanks require an office
building, lots of salary money, and much computer time. MBL just happens to be a bioscience
think tank that instead requires a small fleet of fishing boats and a battalion of librarians.
WHAT KINDS OF TRANSFORMATIONS take place in brains? Perhaps they will help us
understand the brain's versions of blueprints and libraries and computing. The sensory-processing examples are the best known transformations: While vision's lateral inhibition is
perhaps the best-studied, the same principles are seen in skin sensation and hearing. Most
exhibit a version of those Mexican hat arrangements where one region is excitatory but a
surrounding wider region is inhibitory, leading to an optimal spot size -- any larger and the cell
becomes uninterested.
In the moonlight, such inhibition is turned off in some mammalian eyes (certainly cats and
probably ours as well) to increase sensitivity -- another one of the things that Stephen Kuffler
discovered. After our eyes adapt to darkness, big spots of light are even more effective than the
formerly optimal-sized spots -- the cell can no longer tell what size a spot is. Which is why you
can no longer read anything with a less-than-headline-sized typeface in the moonlight. After all,
in such dim light, a photoreceptor has to give a detectible response to a single photon -- whereas
in daylight it is bombarded by a million times as many. Inhibitory surrounds are one way of
regulating sensitivity over that millionfold range.
Originally we called this "surround inhibition." But then those inverted Mexican hat cells were
discovered with excitatory surrounds and inhibitory centers -- so we began to talk instead about
"center-surround antagonism," or simply "lateral inhibition." Whatever you call it, it's seen at
virtually every level of the visual system, from retina to secondary cortical areas. The skin senses
use it. Hearing uses it; a cell maximally sensitive to middle C may be inhibited by tones a half-octave above or below. Lateral inhibition's contrast-enhancing transformations require a broad
wiring, each cell receiving excitation from a wide area, and inhibition from an even wider area.
Or vice versa.
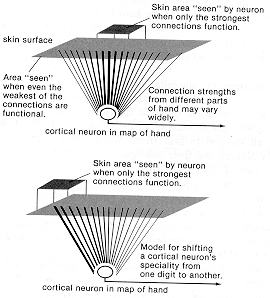
So is this related to the "everything is connected to almost everything" wiring of prenatal
development, with pruning of far-flung connections used to narrow the connections down to a
cone? Probably. Certainly the basic center-surround architecture is present by birth in the
primate visual system. For at least four stages of processing, each cell's view of the world is
from a funnel of converging inputs. The same thing is true of skin sensation: a funnel of
excitation, a wider funnel of inhibition. In theory, this should mean that the higher-order cells
back in the brain receive from ever-widening areas of skin surface, and potentially can respond to
half the body surface (if all the inhibitory surrounds are turned off at each stage!).
REMEMBER THAT CORTICAL MAPS are maps based on the estimated centers of cortical-cell receptive fields, not on their total size (much less their potential size!). If all the inhibitory
surrounds are working full strength, receptive-field centers will seem small, and a center point
easy to define. And so it won't be too much of an exaggeration to say that there is a point-to-point correspondence of the skin (or retinal) surface to the cortical surface -- that we may, in
short, talk meaningfully of a cortical "map" of the sensory surface. But if all the inhibition were
turned off, the "map" might be pretty hard to detect because of the gross smear of anatomical
connections, almost half of the total sensory surface potentially converging upon each cortical
cell.
This widespread anatomical basis for the much narrower functional specialization is the reason
why a cortical cell can shift function, coming to specialize in a different finger than it formerly
did. Everyone thought that cortical maps were pretty fixed -- maybe they are different in
different individuals, but that they were fixed during the lifetime of an individual. But in the
1980's, we were all shocked to hear (from Michael Merzenich, Jon Kaas, Randy Nelson, and
their colleagues) that somatosensory cortical maps were a day-to-day affair, changing size if the
hand was exercised more; if a particular fingertip was regularly rubbed on something (say, a
casino croupier always fingering the deck of cards with his forefinger), more cells in the somatosensory cortex would come to specialize in that finger. And conversely, the size of the average
receptive-field center for a cell specializing in that finger would become smaller.
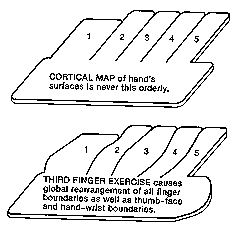
Usually when this happened, the new forefinger cells would come from cells that formerly
specialized in adjacent fingers -- but sometimes from cells that formerly specialized in the face!
The face's connections to such versatile cells were turned down to nothing, while the forefinger's
connections were enhanced -- and so a "retrained worker"!
But they also noticed that some changes in cortical boundaries seemed to occur spontaneously
from week to week, even though the monkey wasn't being trained and was just moving about his
cage. For example, the boundary between face and hand cells in cortex moved from week to
week, back and forth -- some weeks, the cells near the boundary were face specialists, other
weeks the very same cells were thumb specialists. To neurophysiologists, this was
approximately as if you had told us that the state line between California and Oregon was moving
a few miles back and forth from week to week for no apparent reason.
When the researchers trained monkeys to hold one forefinger against a vibrating surface, they
observed threefold increases in the number of cortical cells specializing in the tip of that
forefinger. But the other finger boundaries shifted as well. When "California" was overly
exercised, the California-Oregon boundary shifted -- but so did the Oregon-Washington and
Washington-Canada boundaries! The whole hand map rearranged itself to accommodate the
fingertip exercise; the expansion wasn't just at the expense of the immediate neighbors in the
usual boundary-dispute manner. Furthermore, it wasn't just the other fingers that were squeezed;
much of the expansion was at the expense of the face and wrist representations, as the total
"hand" enlarged. Historical trends may have remade the map of Europe over the centuries, but
whatever remakes cortical maps can be much quicker. One almost has to think of maps as
ephemeral, about as permanent as the arrangement of papers atop my desk.
So learning to play the piano probably does remake your brain, in a very real sense. But so too
might almost any other activity that repeatedly stimulates a hand (or, presumably, foot). I told an
anthropologist friend about this, because she always goes barefoot -- her feet receive much more
detailed sensation that way than when encased in a shoe. Does that increase her foot
representation at the expense of other body parts? Does it decrease her other sensory abilities?
No one knows yet. Is this plasticity why many stroke victims get much better in the weeks and
months after their strokes, with uninjured regions of cortex taking over the jobs originally done
by the injured cortex? Is this why blind people can hear more acutely? Tune in next year.
CORTICAL MAPS ARE EPIPHENOMENA anyway, since they serve no known purpose other
than as guides to the neurophysiologist in the placement of recording electrodes. It is the nerve
cells themselves, like the industries of a city, that are the functional pieces; while street maps do
serve a function for strangers to the city, I cannot think of a comparable function served by the
brain maps we produce. What is so interesting about these ephemeral maps is that they indicate
that there is a lot of retraining of workers in the brain, that the number of cells assigned to a task
(such as analyzing sensation from the forefinger) can be modified on a week-to-week (and
probably a day-to-day) basis.
And most of the plasticity seems to be at the cortical level; the maps of the fingers in the
monkey's thalamus, the relay station just before cortex, are not altered in a similarly dramatic
way. The thalamic nerve cells specializing in the forefinger, however, seem to send axon
branches all over the hand region of somatosensory cortex, not just to the forefinger's current
patch of cortex. This is what probably allows the rapid retraining of a cortical cell: It just
switches from suppressing everything except forefinger to suppressing everything except thumb.
Those cells that switch back and forth between thumb and face presumably have anatomical
connections from both, with one or the other set suppressed. The alternative explanation, that the
thalamic axons sprout new connections and that the old synapses disconnect, is not ruled out --
but the rapidity of the changes is faster than such sprouting processes usually happen in the
nerves to muscle.
Dynamic remapping, going on all the time, suggests that there is an ongoing competition of some
sort that results in the work getting spread around to the available workers. Rather than a
lifetime structure, it may be more like a free-wheeling economy -- perhaps a certain rate of neural
unemployment is used to make sure niches are explored and filled. Are there monopolistic
practices, used to seize power and keep down the newcomers? Is there some central direction, a
circulating hormone that functions like a five-year plan to steer migrant workers in some
directions more than others?
And while economic analogies are perhaps more familiar, it seems likely that the true analogies
are going to be to things even more primitive than economics: Self-organization, darwinian
evolution, and ecosystems. The familiar computer analogies to which we retreat when seeking
an analogy for brain functioning seem totally inappropriate: Computers have a memory that is
kept separate from the processor; they do what they are told to do, rather than seeking out new
niches.
Ever since Darwin and Wallace, we've known that we need to understand evolutionary principles
in order to understand how we came to be, that long road from monkey to ape to hominid to
human. For almost as long, the analogies between ontogeny and phylogeny have made us aware
that darwinian-like processes (such as all that prenatal cell death) are a major part of getting from
fertilized ovum to an adult. But now it looks as if anyone who wants to understand day-to-day
brain functioning (the nature of perception and cognition, the basis of memory, the organization
of behavior) had also better bone up on Darwin.
What we call a mind is nothing but a heap or collection of different perceptions,
united together by certain relations and suppos'd, tho' falsely, to be endow'd with
a perfect simplicity and identity.
the Scottish philosopher David Hume (1711-1776)
Philosophy has succeeded, not without a struggle, in freeing itself from its
obsession with the soul, only to find itself landed with something still more
mysterious and captivating: the fact of man's bodiliness.
the German philosopher Friedrich Nietzsche (1844-1900)