3
FINDING A FAST TRACK TO THE BIG BRAIN:
How Climate Pumps Up Complexity
D'oů venons nous?
Que sommes nous?
Oů allons nous?
Where have we come from?
What are we?
Where are we going?
Paul Gauguin, 1897
The old
joke goes, "But you can't get there from here!" (This was the
response given by the laconic farmer to the city motorist, who
was lost in the back country and asking for directions.) I'm
reminded of it because of our attempts to find a certain
restaurant in the hills near Lake Balaton. We asked questions
-- in German, translated from American English -- of
Hungarian pedestrians and eventually discovered that we had
to drive back downhill to the lakeshore and then take a
different road back up into the hills.
But, of course, you can get there from almost
anywhere (when in the company of astronomers, one has make
allowances for the improbability of ever escaping from a black
hole). It's just that the path may be a little roundabout,
requiring a lot of detailed description about backtracking to
some other junction, rather than a simple "That-a-way."
Progress sometimes requires a temporary dose of
regress. The evolutionary path from an apelike
ancestor to Homo sapiens also requires a lot of
detailed description, including backing up a few times.
Indeed, if intelligence is not one of those much-sought
"general principles" of the universe, the details of the path are
all-important.
Yet self-congratulatory generalizations are about all
we've got in the way of explanations for hominid evolution --
such as "Man the Toolmaker" or the "bigger is smarter is
better" pseudoexplanation of why brains enlarged fourfold.
What we need is a good idea about each leg of the journey to
humanity, the opportunities and provisions along the way, the
hazards and how our ancestors coped with them, plus some
notion of how we avoided drifting back to where we started.
And we've got a lot more to explain than mere cleverness or
brain size -- for instance, ethics, art, music, compassion.
We've come a long way from the apes. There is no
animal currently around that can serve as a suitable stand-in
for our common ancestor with the apes. But, if you imagine
the common ancestor as a composite of the chimpanzee and
gorilla, you're not likely to be far off. While all ape species
have surely changed as well during the last 5-10 million years,
they haven't made the major transitions that separate us from
our common ancestor: walking upright, concealed ovulation,
elaborate language, extensive tool use, accurate throwing for
hunting, and the big brain.
Attention has naturally focussed on how we evolved
from an ape level of language and intelligence to that exhibited
by the remaining hunter-gatherer bands of today -- and on the
acquisition of a fourfold larger brain along the way. Did
toolmaking drive the brain boom, as anthropologists once
proposed? The usefulness of language, as the linguists
propose? The psychologists are naturally in favor of
intelligence as the raison d'être. And at least one
neurophysiologist thinks that it is mostly due to the brain-muscle coordination needed for hunting with projectiles
(although I'm not a sports fan, we neurophysiologists are
fascinated with rapid movements of all kinds). If you asked
a reproductive biologist, there would surely be a key role for
concealed ovulation (no more estrus behaviors advertising the
time of maximum fertility, promoting pair-bonding but also
social cleverness). All of us could be right. Unlike the tale of
the blind men and the elephant, there is more than one right
answer -- because everything in biology has multiple "causes."
There may be multiple ways to be "right" but there are
even more ways to be wrong. And eliminating incorrect
explanations is a key way in which science progresses in many
fields. As the economist Kenneth Boulding once said,
I have revised some folk wisdom lately; one of my
edited proverbs is "Nothing fails like success,"
because you do not learn anything from it. The only
thing we ever learn from is failure. Success only
confirms our superstitions.
For some strange reason which I do not
understand at all a small subculture arose in
Western Europe which legitimated failure.
Science is the only subculture in which failure is
legitimate.
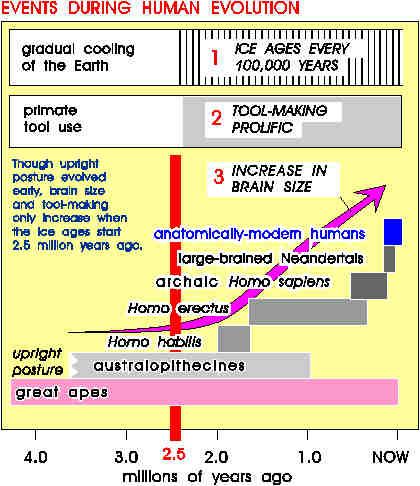
For example, it was once thought -- quite reasonably, I
might add -- that upright posture was caused by the need to
"free up" the hands for toolmaking and, in addition, that a
bigger brain was required for manual dexterity. Thus
toolmaking should precede upright posture and the brain boom,
and parallel their changes. The sequence was even embedded
into popular thought by the opening scene of the movie
2001. Now, thanks to a lot of hard work in the hot
sun by the paleoanthropologists and archaeologists and
geologists, we know that, instead, upright posture
preceded prolific toolmaking by several million years.
Some fossil footprints of a bipedal hominid dated to
3.5 million years ago are virtually identical to those of present-day South American Indians that habitually go barefoot.
Though the facts eliminated that hypothesis for upright
posture, they unfortunately didn't explain what "caused" the
posture to shift to upright.
And we now know that about 2.5 million years ago,
three major trends started up, all at about the same time:
prolific toolmaking, brain enlargement, and the ice ages. Was
one the "cause" of the other two? I can't imagine how
anything done by a hominid could have affected the ice ages
(at least back then -- we surely can cause an ice age now,
should byproducts of our technology increase the marine cloud
cover enough). So that reduces the possibilities to:
1) toolmaking-to-encephalization,
2) encephalization-to-toolmaking,
3) iceage-to-toolmaking, and
4) iceage-to-encephalization
if one ignores such other issues as language (and the null
hypothesis: That they all happened independently of one
another!).
Some crucial archaeological evidence is now available
concerning the first possibility. While brain enlargement
accelerated to achieve the fourfold mark by about 0.1 million
years ago in Homo sapiens, toolmaking had a more
fitful course. From about 1.5 until about 0.3 million years
ago, the brain size of our ancestors doubled -- but toolmaking
suffered from a lack of major developments; the Acheulean
toolkit stayed about the same. So it becomes hard to argue
that innovative toolmaking was what rewarded any bigger-brain variants in the genome. So eliminate #1.
That encephalization might have eventually facilitated
toolmaking (#2) will surprise no one, but it seems a very slow
path to me. That the ice ages might provide a stimulus to
toolmaking (#3) is similarly possible but slow, and the
argument is not as sound as one initially supposes. So did the
ice ages drive the brain boom (#4), which secondarily
facilitated toolmaking? That's still a very interesting
proposition -- indeed, what the rest of this book will examine.
This isn't some general evolutionary principle at work: that's
because the ice ages do not seem to have affected other
mammalian brains in a similar way. One has to examine this
possibly unique journey in great detail, looking for connections
between Pleistocene ecological opportunities and the hominid
skills not shared with the apes. And while concentrating on
"progress," one must remember to look around for
opportunities conferred by regression as well.
THE CLOSER WE LOOK at apes in the wild, the more
we notice that we share many features of their social lives.
The African apes are clever, both in manipulating tools and in
social manipulations of each other -- including both
cooperation and deception, both aggression and peacemaking.
Patiently trained to use sign language in the laboratory, apes
exhibit the ability to use language with about the complexity
shown by a two-year-old child -- a vocabulary of a few
hundred words and simple sentences that make no demands on
syntax -- though in the wild, the apes use only a few dozen
exclamations and a variety of body postures and facial
expressions. That shared body language and gesture is still
used by humans -- when language fails (as by Americans in
Hungary) or just redundantly (as by Italians in Italy). We
have greatly elaborated communications with our serial-sequential languages -- but just how important might that have
been for finding food, defending against raiders?
What about some of the ecological-niche candidates
for shaping up modern man? More than a million years ago,
hominids learned to live outside the tropics and subtropics,
which means that they could cope with winter. This required
some new skills. Clothing for insulation, fire-making, and
shelter would take on considerable importance. Yet none
would seem to require much more intelligence than our ape
cousins demonstrate -- more patience, perhaps, but not much
more cleverness.
The big problem with living through the winter is
food: the choice becomes very restricted for a few months
each year because the gathering is so thin, plants having shed
their edible leaves, and the snow masks what's left for the
taking. To get yourself, much less dependent offspring,
through the winter usually means one of two strategies (though
there are some exceptions): hoarding a surplus from the
summer, or being able to eat grass, which remains nutritious
through the winter, whether bundled as "hay" or dormant on
the ground. The grazing animals manage to find enough grass
beneath the snow, and they have evolved the teeth and
digestive enzymes they need to utilize it. Humans haven't, but
they have managed to eat animals that eat grass by becoming
skillful enough at hunting.
Hoarding sometimes works, but usually only with
hunting as a backup: sometimes the harvest is insufficient,
sometimes the rats break into your grain hoard, or the wolves
discover the frozen carcass and gobble up the meat on which
you were relying to get through the last half of winter. Living
in the temperate zones tends to suggest that hominid hunting
techniques had become more reliable than those opportunistic
snatch-and-grab tactics used by chimpanzees.
But what do hunting skills have to do with our more
valued aspects of humanity: language, consciousness, ethics,
music? Actually, thanks to the neural machinery needed for
accurate throwing, quite a lot -- because all of the afore-mentioned happen to be aspects of the serial-order behaviors
in which the brain's left hemisphere specializes. Perhaps,
when not throwing projectiles or swinging a club, the neural
sequencing machinery can be used for speaking a sentence, or
planning for tomorrow, or feeling dismay when seeing a
tragedy unfold (or laugh at the surprise ending, counter to your
expectations). Some such neural machinery is secondarily
useful for composing a melody, or playing chess, or dancing,
or kicking. Spare-time use of machinery in the off-hours for
secondary uses didn't start with playing games on the office
computer: conversion of function is a major mechanism of
evolution, described by Charles Darwin in the middle of the
nineteenth century. Natural selection acting on any
one serial-order skill might tend to improve all
of them, just because they all utilize the same neural
machinery for creating novel sequences of muscle commands.
WHICH WAY TO OUR BIG BRAIN? Given all the
advantages of being smart, there is surely more than one way
to become intelligent. Indeed, those who have thought about
the SETI problem often observe that "Surely X would
be useful." And the theorist of evolutionary intelligence can
seldom rule out that route, in the manner that a physicist
manages to rule out 99 percent of the theories offered to
explain a phenomenon, just showing that X would also
cause Y to happen -- and it hasn't, therefore rule that
one out too. In biology, there are always multiple causes of
everything we study, so we can't have the physicist's kind of
confidence in our attempts to eliminate causative factors.
But one way to make progress in clearing away the
minor causes and concentrating on the major movers is to ask,
"Yes, but how fast would such a change happen?" In biology,
fast tracks tend to preempt slow tracks. And sometimes,
evolution happens so rapidly that only a fast track could have
done it.
Our big brains are a prime example of rapid evolution:
they have increased fourfold in size in a mere 2.5 million
years. That's almost unbelievably fast by the standards of
evolutionary biology. The rapidity itself is a clue as to the
evolutionary causes of brain size increase. It tends to rule
out slow tracks such as smarter-is-better, where the
incremental payoff for each ten percent increase is small
indeed.
So what controls evolutionary rates? Some factors
are: Producing individual variations (via mutations and
permutations). The severity of selection (droughts every
decade). The rate of evolutionary inventions (those
conversions of function, the compounding of mechanisms).
The invention of a new niche, or finding an empty one, makes
for boom times (of which, more later). The prevention of
backsliding (via reproductive ratchets and similar stabilities).
And more.
The hominid's Great Encephalization is so rapid that
one tends to look for scenarios that incorporate a number of
these factors. We may not have used every possible way of
speeding up our brain changes, but we likely used quite a few
of them.
TO SHOW HOW SELECTION shapes up populations,
short of writing a textbook on all these factors, one can try
telling a bedtime story about a bear. Bears are popular here in
central Europe; they're "emblematic," as the anthropologists
might say. My exemplar bear will henceforth be known as
Mama Bear.
At the opening of Act I, we see Mama Bear and her
two baby bears, gambolling about in the summer sunshine. So
ends Act I (and the traditional version of the bedtime story; the
following might be considered a revisionist adult version).
Act II is a year later. Mama Bear weaned the two
baby bears after a half-year of suckling them. Then after
putting up with them for a winter of hibernation, she kicked
them out into the real world to fend for themselves. This is
Mama Bear's summer for getting pregnant again; next winter
she will again give birth to another pair.
Act III is another two years later, with another two
half-grown bears let loose on the world. Mama Bear may do
this five or ten times during her lifetime, if she stays healthy.
Unfortunately, a little arithmetic shows that this story
doesn't have a happy ending. How many bears can the
environment feed? Obviously, that's the average bear
population. And that means, on average, only two babies per
mother get to grow up and become a parent, out of the dozen
or two that she produces. The maximum population level is
not set by the birth rate but by the number of job slots
afforded by the environmental niche occupied by bears. And
that is a complicated function of food availability, suitable
nesting sites, predator populations, pathogens, parasites, and
such. Only in boom times does birth rate have much to do
with it.
That means the average Mama Bear is raising five-to-ten times more baby bears than can possibly survive, absent,
of course, miracles -- approximately what one might call it
when a new niche happens to open up, either by new territory
becoming available (as when the Alaskan brown bear
discovered the ice-free corridor, at the same time as the Paleo-Indians) or when a new way of making a living is discovered
(as when bears learned to go fishing in streams for salmon
migrating upstream). For cats and dogs, the waste is even
worse: say, six per litter, and five or ten litters per mother,
but only enough adult food for two of them to survive. I
know one cat, living in optimal circumstances, who has given
birth to more than 200 offspring, a hundred times her quota.
So again (at least in nature) there are a lot of animals that are
going to die of starvation, becoming food for a predator.
Why does Mama Bear spend all that effort raising
many times more offspring than the market will bear (no pun
intended)? It's called "keeping up with the Joneses." For
Mama Bear to have a gene Q for doing less would
result in fewer such gene Q's in the next generation;
this gene would "work itself out of business." Indeed, this
situation is like an arms race; if a variant arises that can
successfully raise triplets, her genes will soon take over the
population, replacing the two-at-a-time genes, making triplets
the new standard, and so on to even bigger litters.
If triplets don't happen now, it's probably only because
each would be so stunted in size from crowding in
utero as to be less competitive; three scrawny ones
being worse than two of the standard twin-size. The bigger
the litter, the higher the percentage of manufacturing defects --
and the less postnatal parental attention per offspring. In
boom times, all offspring might survive -- and indeed both
bears and humans shift to larger litters when resources are
bountiful.
This is an Alice-in-Wonderland sort of principle: the
Red Queen told Alice that you have to keep running just to
stay in the same place. The Red Queen is one of the reasons
we talk of a naturalist fallacy. "What's natural is good," is,
alas, another example of substituting upbeat wishful thinking
for familiarizing oneself with the available evidence. Across
many species of mammals, the amount of effort devoted to
reproduction bears little relationship to niche size. Nature just
hasn't developed a way of limiting overproduction of baby
bears (or any other species, although humans may yet become
an exception), and most animals spend nearly all their life just
raising youngsters with which to feed the predators and
pathogens.
THIS UNHAPPY STATE OF AFFAIRS (by humane
standards) provides, however, the raw material on which
natural selection operates. Though we usually focus our
attention on the adult population, it's really the young (after
their parents cease to care for them) that are the prime objects
of natural selection. The young are where the action is. The
young are comparatively inexperienced in critical areas -- and
there are many more of them, compared to adults.
While predators also cull the old, that's not an example
of natural selection at work: whatever genes the older animals
are going to get into the next generation is determined earlier
in their life, so whether they die now or later isn't going to
shape up the ongoing population of genes. This is why there
hasn't been much natural selection against late-developing
aspects of gene repertoires, such as Huntington's chorea, or
cerebrovascular disease, or Alzheimer's senility, or the inability
to digest milk that sometimes develops in the midthirties.
Predators also cull the sick. That, however, promotes
natural selection for the immune system's capabilities. Natural
selection doesn't just work through predators and food
availability but also via childhood diseases: it takes years to
build up immunity to the common diseases, which is why
older workers have fewer respiratory infections than younger
workers. In one sense, you actually become healthier as you
grow older!
But in the real world of Darwinism, if an animal
becomes weak from a virus, a predator eats it. If the animal
becomes weak from inability to find food, a predator
dispatches it. The young are comparatively inexperienced in
both areas. Lacking a change in the average environment (of
which, more later), the top 10-to-20 percent of the young bears
survive -- which is quite a shaping-up. Some of the others
make it too by accident, and some of the top ones die by being
in the wrong place at the wrong time, as when struck by
lightning.
Droughts make selection even more severe, but the
overproduction among mammals causes a lot of severe natural
selection, just as a baseline. Somehow, I doubt that more
severe selection (e.g., harder winters during the ice ages) was
the cause of the Great Encephalization, especially given that
other animal's brains didn't similarly enlarge during the ice
ages.
EVEN THE BABY BEARS that escape natural selection
for a normal adult lifespan may still have a big problem.
Another form of selection operates because not all get to
breed.
In most animals, nearly all surviving females get to
bear offspring (there are some exceptions among social insects
and dog packs, where a dominant female may inhibit the
reproduction of subordinate females). In many types of mating
systems, however, quite a few males are locked out of
propagating their genes. Harems are the most obvious
example.
What determines which males get their genes into the
next generation? Sometimes, brute force decides: head-to-head competitions between males for control of a harem,
where male body size and armaments count heavily. In some
other mating systems, there is female choice, typically for
healthy-looking males. This potentially augments the tendency
of natural selection to promote improvements in body styles.
But health isn't always judged by something truly relevant,
such as having the prospective suitors run a marathon or
collect a week's food for a family. There is usually some
substitute indicator of health used, some stand-in -- perhaps
agility in a mating dance, or the condition of a male bird's
plumage.
And this can lead to appearance mattering more than
reality, with some cosmetic trimmings all-important. If shiny
plumage is the criterion by which a female bird selects one
male over another, you can see an arms race in plumage, such
as the iridescent peacock tails. Sometimes it is feather length
-- and so you see some absurdly long tails in species such as
the bower birds and magpies.
So sexual selection is based, not on the
elements of the natural environment such as food availability,
predators, pathogens, nesting sites (those are the elements of
natural selection, though it might be better called
environmental selection) but on reproductive
peculiarities, many of which no longer function in any
reasonable way. Those absurdly long tails may impede flying
abilities and those bright feathers tend to give away one's
location to predators (and so sexual selection may conflict with
natural selection, balancing each other out). Male gorillas are
so heavy as adults that they cannot take to the trees when a
predator approaches, in the manner of the adult females and
adolescent males -- they have to stay and fight! One presumes
that some such counterbalancing with natural selection is why
sexual selection doesn't often keep proceeding to absurd
lengths.
WHAT CONTROLS EVOLUTIONARY RATES, and so
the length of time it takes to shape up a new feature? Most
people would immediately suggest mutation rate, how fast the
cosmic rays or mutagenic chemicals can introduce errors into
the DNA strings. While an extra dose of radiation can indeed
augment variability in offspring, gene permutation is
probably the most important aspect, that shuffling of the
chromosomes that takes place during crossing-over as new
sperm and ova are made. From generation to generation, far
more variability in offspring is created by permutations than by
new mutations.
Furthermore, evolution above the one-cell level didn't
really get going until crossing-over was invented by eukaryotes
about one billion years ago; promptly thereafter, multicellular
life developed in a big way, inventing about 50 major ways of
structuring a body plan during the next half-billion years.
Mutation didn't accomplish that: it was permutation. What
affects the rates at which genes come to stick together, or
develop new points at which to break apart during crossing-over? That's one of the unsolved problems of basic biology.
Among other factors, the reproductive arms race and
its wastage must partially control the opportunity for natural
selection to act on the variants thrown up by gene shuffling
and mutation: everything else being equal, cats ought to
evolve faster than bears because they overproduce more (their
top 5 percent might survive, rather than the bear's top 10-20
percent). But fortunately we can avoid discussing the "cannon
fodder" principle ("the more waste, the faster we evolve!"),
because climate is the most obvious variable when it comes to
fast vs. slow evolution.
The most rapid of environmental cycles are the daily
ones associated with day-night and with the tides. Any planet
is going to have solar tides, so long as it has oceans and
doesn't keep one face always towards its gravitational attractor.
If a planet has two attractors (as does ours), that's even better
for speeding evolution. Thanks to the moon's tidal forces
adding to (and then, for half a month, opposing) the sun's
gravity, there are also monthly and yearly cycles of extreme
low tides. The tides serve to select for intertidal plants and
animals that can survive in a second kind of environment for
longer and longer intervals -- perhaps until becoming land-dwellers.
WHILE LAKE BALATON is Hungary's largest body of
water, the tides here are about as conspicuous as the
Hungarian Navy. One sees only wind-driven waves of greater
and lesser proportions, keeping the shoreline wet. But it
reminds me of the shorelines back home in Puget Sound where
the sea level varies each day over an average range of more
than one story high, and so a lot of beach alternates between
being underwater and being temporarily above water, drying
out in the sunshine.
At some such intertidal zone of 450 million years ago,
life came ashore. A species exposed to the monthly low tide
series was undergoing natural selection for mechanisms that
would keep it going in two different environments, both free-flowing water and up in the air. For the intertidal species, the
tides provided daily waves of selection for the abilities needed
to survive extreme variations in moisture, pH, salinity,
oxygenation, and temperature. Had such tidal selection instead
happened once per century, the fanciest land animal these days
might be a floundering lungfish.
It takes a swamp-and-tide-flat zoologist to tell
you about life; it is in this domain that the living
suffer great extremes, it is here that the water-failures, driven to desperation, make starts in a
new element. It is here that strange compromises
are made and new senses are born.... [In] the
mangrove swamps by the Niger, fish climb trees
and ogle uneasy naturalists who try unsuccessfully
to chase them back into the water. There are
things still coming ashore.
Loren Eiseley, The Immense
Journey, 1957.
ONCE ASHORE, there are some yearly variations in
environment outside the tropics -- better known as the seasons.
Thanks to axial tilt and land surface in the temperate zone
(mostly Northern Hemisphere these days, the tip of South
America excepted), we have had yearly cycles of selection for
species able to survive both summer and winter weather (most
species simply stick to the tropics).
But for those who do evolve the mechanisms to endure
both winter and summer extremes, there will be yearly waves
of selection, operating upon that huge overproduction of the
Mama Bears of the temperate zone. While not as frequent as
the daily and monthly cycles of the tides, wintertime selection
cycles might cause more rapid evolution in the temperate
zones than in the tropics -- at least for winter-related body
features. And for the behavioral traits needed in the
wintertime (predation skills are particularly important in many
animal species, as plant life becomes dormant).
THANKS TO CYCLES in the atmosphere-ocean system,
we have multiyear cycles of drought. Somewhat understood
are the monsoon variations in the Indian Ocean, and El Nińo's
twice-per-decade cycle in the weather systems of the Pacific
Ocean. Among human populations, the families of South
American fishermen are most affected -- but the bird
populations of many Pacific islands crash to ten percent of
their usual numbers. Recently, some of the U.S. midwestern
droughts have been linked to El Nińo as well.
Pathogens also have multiyear cycles, as in the
shellfish population crashes. Forest fires occur every few
decades and, near a shoreline or watercourse, floods occur
several times each century (if not more often). Less
systematic are the meteor strikes and volcanos that darken the
atmosphere (though some of the more famous examples of
post-volcanic cooling, such as the year-without-a-summer in
1816, may yet turn out to be unrecognized visitations of El
Nińo).
Those trying to live on the margins of a habitat are the
hardest hit. In Europe, most traces of people who lived at low
population densities have been lost by the reuse of sites by the
peoples that followed. But in the New World, one can do
better: many "stone age" sites have been discovered relatively
intact. For example, in the U.S. Southwest, rainfall improved
about A.D. 1050 and many new Anasazi villages sprung up all
over the area; by 1130, they were all abandoned and even the
major population centers were dwindling. So the archaeologist
gets to see a window in time, largely uncontaminated by what
followed. Boom and bust is common in nature, not just in
economies.
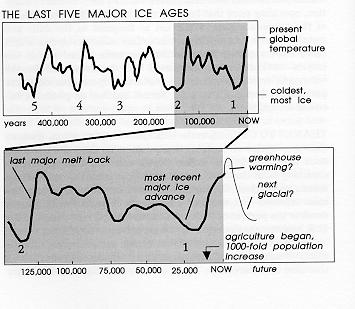
THANKS TO VARIATIONS in the earth's axial tilt and
the drift in the season when perihelion is reached, we've had
100,000-year-long major climatic cycles to shrink and expand
the temperate zone populations. Of which, more later.
There is also lots of back and forth within each ice
age, with perhaps five minor retreats in the ice sheets between
each major meltback. Back and forth. So that means a major
climate change about every 10,000 years (and within just the
last 120,000 years, "cold spikes" have also occurred about
every 6,000 years in the North Atlantic region alone).
And we know that there are centuries-long fluctuations
within this as well. The Little Ice Age was a period of cooler
climate between about 1200 and 1800, made even worse on
occasion by some volcanic eruptions that clouded the
atmosphere and cooled the weather for several years at a time.
[There] are two ways in which a
creature can seek to survive in a jungle
environment. One way [known as
wedging] is to compete fiercely and
successfully for an existing niche with
other creatures that are trying to occupy
it. The other way is to find a wholly
unoccupied niche....
Herbert A. Simon, 1983
MASS EXTINCTIONS also affect evolutionary rates,
thanks to whatever happens every 28 million years (volcanos,
asteroids, and meteors are possible, but some favor comets
since a scheme has been suggested whereby the recurrences
would cycle every 28 million years). After each mass
extinction, wherever its cause turns out to be, there are
opportunities for new species to fill vacated niches. This is an
extreme case of boom and bust, and so it is worth examining
it in more detail.
Darwin realized that evolution could be slow if
efficiency was the only factor. An improving species (say, one
better able to utilize a particular food) would have to wedge its
way into the niche of another species already utilizing the
resource. This metaphor of the wedge is very useful, but some
of the major advances in evolution occur when no wedging is
needed -- because of the empty niches. Some empty niches
are just there, waiting to be found: There are no woodpeckers
on New Guinea despite dozens of species of woodpeckers on
Borneo and Sumatra in similar forests. And filled niches can
be emptied, as occurs in an extinction.
In periods of rapid diversification, a whole series of
empty niches are discovered -- as when some uninhabited
islands are discovered, as has happened on both land and sea
after the mass extinctions of 250 million years ago and 65
million years ago (those were merely the biggest extinctions;
minor ones seem to happen about every 28 million years, the
last one about 10-11 million years ago). After the dust clears,
competition within a species is not important for awhile; nearly
all the offspring get to live and reproduce, allowing for
variations that would ordinarily be culled by natural selection
to survive and, indeed, be elaborated in successive generations.
And so body styles vary widely; when the niches start to fill
up, many will be eliminated and a more standardized model
will take over.
But some variants may become associated with a
stable niche. If they no longer interbreed with the parent
species, as often happens in such situations, then they will
constitute a new species. This serves as a ratchet, preventing
backsliding to some average body style. Much evolution, in
the sense of change in body form, has been temporary because
it did not find a new stable point from which backsliding was
prevented. Speciation, i.e., inability to interbreed effectively
or reproductive isolation, is the prime (though
not the only) means by which stratified stability occurs.
BUT BEFORE SPECIATION OCCURS, isolation has
usually happened by more temporary means. Because the
environment is often patchy, there are subpopulations of a
species that mostly interbreed within their group. They could
interbreed between groups, given the opportunity, but they
don't -- usually because of some geographical barrier such as
a mountain range, or a wide expanse of territory lacking their
kind of food resources. And so the members of a species are
often found in dozens of relatively independent colonies: call
the effectively interbreeding group a deme if you like,
though population or subpopulation or
colony will also suffice.
When natural selection has been episodically severe,
a population may be completely eliminated and the territory's
resources may go untapped until straying members of another
population discover it -- and have themselves a population
explosion, founding a new deme.
Consider, however, what happens in less severe
population crashes. Anyone who regularly visits a natural
setting from year to year, as I do at the Friday Harbor Labs's
nature preserve in the San Juan Islands, must be impressed
with the yearly fluctuations in the number of common species
such as deer and rabbit. Some years, there will be a dozen
rabbits outside the front door of the main labs, mowing the
lawn; other years, you're lucky to see two rabbits when
walking around the entire several acres of lab buildings and
housing units (and the maintenance personnel have to crank up
the lawnmowers that year). Some years, there will be a dozen
deer grazing amidst the buildings; other years, none are seen
for days at a time. Some years, the raccoons are particularly
prominent.
These fluctuations are not due to variations in food
resources, nor nesting sites. Some are due to hard winters.
But most, I suspect, are due to diseases that nearly decimate
the island's population within several years. The survivors are
the few whose genes allowed their immune systems to cope
with the infection. Some of their offspring will inherit those
genes, and so the population may start rebuilding even in the
continued presence of the virus.
Over a century, a population will be exposed to quite
a few similar episodes. If the population is modest in size, its
genes may be severely edited by such episodes, and not just
those for the immune system. Those whose constitutions are
particularly robust will survive illness better, and so the plague
years (as we would call them, were this a human population)
also shape the genome in the direction of a specialty in this
island's environment: its climate, its food resources, etc.
Illness, by weakening the individual, sharpens the importance
of that island's natural environment. Populations on other
islands may be shaped somewhat differently, both by chance
and by the differing environment there.
Boom and bust cycles edit small island populations far
faster than they would a mainland population. You can see
why a Mama Bear that produced triplets rather than twins
would spread her triplet genes around rapidly, given the
occasions where an island's bear population had been depleted
by a virus. On the mainland, the adjacent populations
unaffected by an epidemic could later move into a depleted
area, the merely lucky competing with the offspring of the
survivors who had "earned" the chance. Large mainland
populations buffer rapid change -- and incidentally help insure
against the loss of valuable genes, as can easily happen in a
small island population.
Back in the millennia long past, when humans were
sparsely located over the surface of the earth, each tribe was
effectively on an island. Now evolution is slowed down to a
crawl, simply because there is little isolation anymore. A large
population evolves slowly in comparison to a small deme.
AGRICULTURE GREATLY REDUCED the influence of
natural selection on most humans (while we still die, the
evolutionary issue is who dies when). "Being better"
now shapes up the human gene pool rather slowly. And that's
probably been true on most continents for the last few
millennia, perhaps most of the period since the continental ice
sheets melted about 12,000 years ago. Which is not to say
that evolution ("change") won't occur in the future, only that
traditional darwinian processes will perhaps play a minor role
in guiding it, compared to biotechnology and such
environmental novelties as air pollution.
So the combination of transportation (allowing
different demes to intermarry more easily, which can have the
advantage of hybrid vigor) and agriculture (serving, along with
sanitation and medicine, to allow most variants born to survive
and reproduce, for yet another round of variation) give rise to
a conclusion about future human evolution that is very
different from the Social Darwinism views of the late
nineteenth century and the eugenics views early in this
century. The eugenicists (not, interestingly, most geneticists)
thought that artificial selection was important to improve
human bloodlines, preventing the mentally ill and the epileptic
from reproducing, and encouraging marriages between those
possessing favored traits.
Given the successes of animal breeding, eugenics was
in many ways a perfectly reasonable hypothesis, given how
little was known at the time. I think few people realized how
slowly things worked -- absent that Mama Bear scale of
wastage, absent new niches to explore. Incorporating this
early scientific speculation into German nationalistic slogans
says more about the need for morale-building than it does
about science.
By the time that the Nazi government got hold of the
eugenics idea (or at least its vocabulary), many geneticists and
evolutionary biologists had backed off from any support of the
eugenics movement. Scientifically, concepts were changing.
Geographic isolation leading to reproductive isolation and
speciation, etc., are insights from the thirties and forties,
particularly from mathematical types such as Sewell Wright,
and the early Forties saw the emergence of the Modern
Synthesis of darwinism with genetics. It took a while for this
news to spread. The major German societies of physical
anthropologists indeed collaborated with the Nazi program of
racial hygiene. And in the United States, awareness was not
markedly better: Even in 1939, the American Association of
Physical Anthropologists tabled a proposed resolution
condemning Nazi racial myths (as the 1989 president of that
association, Matt Cartmill, has recently noted in a sobering
book review).
Part of the problem was that news travels surprisingly
slowly between different disciplines -- in this case,
evolutionary biology and anthropology. Even today, I am
surprised at how little anthropologists concerned with hominid
evolution seem to know about evolutionary biology of the
kinds summarized thus far; their major concepts such as the
savannah and scavenging are seldom evaluated in terms of
anything other than slow adaptations, Darwin's original
valuable insight. Fortunately, ignoring fast tracks and isolation
opportunities (or developmental linkage, or compounding, or
conversions of function) has none of the potential for societal
misuse, at least when compared to transplanting animal
breeding considerations into the human sphere.
Ignorance doesn't merely slow science down:
ignorance also leads to mistakes. One of the seldom-realized
benefits of science has been what knowledge has allowed us
to avoid: the quack remedies and their tendency to delay
effective action until too late, the buildings that collapse from
trial-and-error construction methods, the invasion of pests
because of having ignorantly killed off a natural predator. If
history is any guide, our changing concept of human origins
will enable us to avoid some of the problems in education and
health care occasioned by our ignorance of how we humans
function.
EVOLUTIONARY CHANGE is not only more rapid in
small groups, but it is more likely to become "permanent" in
a temporarily isolated setting. That is to say, something may
happen that makes interbreeding less likely, even when the
geographic barrier is removed (as when an ice age's lower sea
level reconnects all of the islands in the San Juan Archipelago
with one another).
Sometimes this is a chromosomal rearrangement in an
island's population; mating between individuals with the
rearranged and regular chromosomal patterns may have a high
rate of spontaneous abortion or, if there are offspring, they
may be sterile and so fail to propagate the lineage. Other
times, there is simply less of a tendency to mate with a
member of an out-group: behavior can effectively keep
descendants of an island's population, now expanded over a
broader landscape, from mating except with one another. The
classic example is when mating seasons have shifted: on a
mountainous island with late-melting snow, the mating season
may have become a month later than usual, as those variants
had offspring which survived better. And so that island's
population, when mixed up with the general population by
lowered sea level, still tend to mate with their original group
for millennia thereafter, simply because the two groups are
never sexually receptive at the same time.
Such prevention of backsliding might be called a
reproductive ratchet. While mere "attractiveness" of physical
appearance contributes to this tendency, culture tends to
augment it: Erik Erikson noted the "excessive" amount of
human energy that preliterate peoples spent in simply being
different from one another. He calls it "pseudospeciation."
Reproductive isolation that is truly persistent is another
way of saying that a new species has been formed: the traits
shaped up by that island have to stick together,
because they are simply unable to mix with those of the parent
population. Various traits may have altered in an island's
population that have nothing to do with reproduction -- but
they too will be protected against the dilution caused by
mixing with the main population. A reproductive ratchet
speeds evolution.
SPECIES TEND TO DIVERGE a bit after reproductive
isolation occurs because of the Exclusion Principle: one
species per niche. If two species tend to make their living in
exactly the same way, are subject to about the same viruses
and parasites, and so forth, then after enough time has passed,
one of those species will probably decline in numbers and
eventually go extinct in the area where the two species' ranges
overlap. They won't be equally adept at utilizing the
resources: one species will be more efficient.
There may, of course, also be some antagonism
between such closely related species: if you see two species
peacefully coexisting, they are probably not competing with
one another for resources or nesting spots, etc. The
antagonism speeds up the decline of one species, but efficiency
is the most fundamental cause of such an extinction. This
competition between species, certainly the usual
nonbiologist's image of natural selection at work, is, however,
fairly infrequent; most competition is within a species,
involving things such as superior child-rearing.
Animals with a broad ecological niche, such as
monkeys that can efficiently eat many different kinds of fruit
but at different seasons, may so exclude more species than a
narrow specialist like the bamboo-eating panda does. And
humans have one of the broadest ecological niches of all, so
it is not surprising that we have few close relatives left (and
all five of the ape species are now endangered by human
activities). We have created a wide swath of exclusion, and
can only lessen the damage by substantial conservation efforts.
When bigger-brained species of prehumans formed on
some island (real or virtual), they were probably capable of
making their living in new ways or of exploiting former
resources more efficiently, e.g., through food preparation
technologies. If they were reproductively isolated from their
parent species, they would tend to take over in local regions
after a few millennia, even without antagonism between the
two groups.
WHEN THE ICE AGES came along, the lowering sea
levels caused a lot of islands to be reconnected with each
other. And melting later caused new islands to form -- not
only literal islands, but some virtual islands as well: those
mountain-top "islands" in the tropics to which the mountain
gorilla is limited (by its need for 60 pounds a day!) tend to
shrink and expand with the climatic changes seen even in the
tropics when the ocean currents rearranged themselves.
And then, of course, there are the geographic barriers
created by the ice sheets themselves. As Ernst Mayr notes,
they are likely to induce speciation:
During the height of the glaciation, the ranges of
many temperate-zone species contracted into small
pockets, so-called glacial refuges, which persisted
south of the area of glaciation. In Europe, for
instance, the Alpine and northern ice caps
approached each other to within 300 miles,
separated by icy wind-swept steppes. The forest
animals retreated into southwestern or
southeastern Europe. When conditions improved
at the end of glaciation and the populations in the
refuges expanded northward, the isolates in
southwestern and southeastern Europe had, in
many cases, become sufficiently distinct from
each other to form hybrid zones in central Europe.
Animals that could survive in the steppes were on small
"islands" indeed. But the animals that needed forest were
pushed toward the Iberian Peninsula and Greece, effectively
divided by the Alps from opportunities to interbreed. These
"islands" were much larger than the ones between the ice
sheets, but apparently still small enough for speciation to
occur. "Pleistocene forest" refugia also occurred in the
Americas to the south of the ice sheets: mountains in Arizona,
for example, were a refuge for temperate-zone species forced
south from Canada.
In fact, Hungary is right near the boundary between
the group of birds associated with the Iberian refuge and the
group associated with the Balkan refuge. The experienced
bird-watchers among us could probably find some hybrids
right around Lake Balaton, crosses between Iberian and Balkan
peninsular species, living evidence of the icy wedge that
disappeared 12,000 years ago.
SO A SPEEDY SCENARIO for hominid encephalization
would likely be set in the temperate zone, where every year
the winter speeds up natural selection. The temperate zone
would provide exposure to the ice age's tendencies to create
islands on which evolutionary change is faster -- and
incidentally enhance speciation opportunities and reduce
backsliding.
The ice ages also provide a lot of empty niches to fill,
simply because they are forever changing the landscape. Near
a glacier, only grasses grow. Farther back, forests get started.
Birds find them quickly, but mammals take a little longer.
Each species that comes upon a big uninhabited area enjoys a
population boom. Sometimes an "adaptive radiation" occurs,
diverse forms arising while the competitive rules are
inoperative. The big brain is expensive (not just in terms of
blood supply, but apparently in terms of a long vulnerable
childhood), and it might have taken a profound dose of "good
times" to allow it to develop, some major new resource
becoming attainable (such as being able to eat meat every day,
rather than once a month).
We tend to emphasize the conditions in Africa when
talking of hominid evolution. The older fossils are found there
and adaptationist theory tends to emphasize local adaptations
to local environments (rather than carryovers from more distant
former environments). One of the minor points that this book
has to make is that ice ages cause temperate zone traits to
become far more important than one might initially think.
Except near a few mountains, cold weather plays little role in
Africa; during an ice age, while equatorial regions may house
glaciers high on volcanos, most species can easily escape
natural selection for cold climate by retreating to a lower
elevation, an option not always available in the temperate zone
(especially Europe, where southern retreat is often limited by
the Mediterranean).
Hominids spread out of Africa, on the present
evidence, about 1.4 million years ago and, as more digs are
conducted in the temperate zone, the dates will likely become
even earlier. Indeed, we are faced with the probability that
after such a date, the African models of hominid may have
been developed elsewhere: that some of their features were
shaped up in the temperate zone, and later spread back into
Africa. That would mean that such "African" hominids had
some temperate zone specializations that weren't really
essential in the tropics. And the big brain may be one of
them.
I JUST LAUGHED, remembering the time when I was
teased about "walking on water." It was in the bottom of the
Grand Canyon, rafting on the Colorado River. Trying to pull
the boats into the beach near Phantom Ranch, we encountered
a large sandbar offshore -- basically a standing wave of sand,
providing a narrow ridgeline just under the waters' surface but
rapidly falling off into over-your-head depths. And so I finally
got out of the boat, stood ankle-deep atop the sandbar, and
pulled the boat down to where there was a break in the
sandbar. Then I sloshed back up the sandbar to help a second
boat that was stuck. One of its occupants couldn't resist:
"Say, Doc, while you're at it, suppose you could change this
canteen of water into some wine?"
This is surely standard repartee among even devoutly
religious fishermen, repeated many times over the centuries.
And so I couldn't help wondering, when I visited the Sea of
Galilee, if there were sandbars. One winter day, I sat there on
the southern shore near its outlet into the River Jordan, eating
a picnic lunch while facing into a cool on-shore breeze,
looking out over the large lake with the snow-covered uplands
of the Golan Heights as its backdrop, remembering the
1.4 million year old traces of Homo erectus
found a few kilometers to the south in the Jordan Valley,
between the Galilee and the Dead Sea -- right on the path out
of the bottleneck from Africa into Asia.
And I remember contemplating the shallow bottom
and the wind-driven waves that often produce sandbars.
Walking across sandbars is, when you know your way around,
a shortcut that saves much time over the long circuitous route
along the shoreline. A hidden standing wave, no less. If
visitors don't know about the submerged sandbar, it must look
pretty strange.
In paleoanthropology, one concentrates on the stones
and bones, hoping that they will yield some clues to function,
what was serving to shape up the new species. But functions
have shortcuts too, hidden structure that supports a new way
of making a living.
�
We have learned all the answers, all the answers:
It is the question that we do not know.
At the moment we are an ignorant species,
flummoxed by the puzzles of who we are, where we
came from, and what we are for. It is a gamble to bet
on science for moving ahead, but it is, in my view, the
only game in town.
Lewis Thomas

 The Ascent of Mind
(Bantam 1990) is my book on the
ice ages and how human intelligence evolved; the
"throwing theory" is one aspect.
The Ascent of Mind
(Bantam 1990) is my book on the
ice ages and how human intelligence evolved; the
"throwing theory" is one aspect.