|
Conversations with Neil’s Brain The Neural Nature of Thought & Language Copyright 1994 by William H. Calvin and George A. Ojemann. You may download this for personal reading but may not redistribute or archive without permission (exception: teachers should feel free to print out a chapter and photocopy it for students). William H. Calvin, Ph.D., is a neurophysiologist on the faculty of the Department of Psychiatry and Behavioral Sciences, University of Washington. George A. Ojemann, M.D., is a neurosurgeon and neurophysiologist on the faculty of the Department of Neurological Surgery, University of Washington. |
|
Tuning Up the Brain by Pruning
NEIL HAD BEEN PRUNING his apple trees, taking out his frustrations (or so he said) with a
little hard labor. Even though the decision to operate had been made, the surgery schedule was
filled for months ahead. But since he lived nearby, there was a chance Neil could take advantage
of another patient’s cancellation on short notice. All it took for elective neurosurgery to
be postponed was someone catching a substantial head cold. |
|
WE EXPLORE OUR WORLD, seeking sensations. Some come to us unbidden, such as the
sound and sight of lightning. Others we seek out by touching or tasting or smelling, just as we
strain to hear a faint sound or move our head and eyes to get a better view. “I’m glad to hear that sensation-seeking is normal,” Neil said. We can ignore the movements and just talk about “sensations” and “inputs” to the brain, but movement is really an integral part of sensation. If forgotten too long, it can lead you into an overly abstract notion of a little person inside that receives the inputs and decides on the outputs. In reality, outputs are being produced at a number of stages of the analysis. But, despite the hazards, let’s stick to the input path for a moment, the passive sensation side of things. The input path always has a series of stages. The interface with the external environment is a sensor that converts some form of energy into the electrical signals that the nervous system uses for comparisons. We’re always comparing. We have sensors buried in our muscles and joints that tell us where our arm is currently located. Although bending a single hair on your wrist will produce a sensation, hairs are seldom bent one at a time. It’s the comparison with neighbors that enables you to discern whether the sensation is produced by a breeze, a pencil point, a shirtsleeve, or a wristwatch. For our skin sensations, this comparison is not done in the skin. Rather, the axons of the sensory neurons carry impulses back into the spinal cord. Neurons there do the comparisons. Or at least they start the job — they are the second neurons in a chain of analysis that extends to a third neuron in the thalamus, a fourth one in the sensory strip of cerebral cortex, and even more in other “association cortex” areas. Of course, it’s not a real chain of neurons but rather a web. 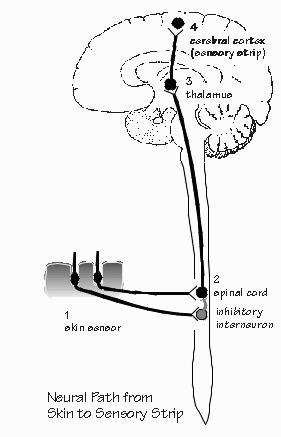 [FIGURE 51 Neural path from skin to sensory strip] “Association cortex? Doesn’t all the cortex make associations?” Indeed, all of the brain and spinal cord. That’s an old name for all the neocortical areas exclusive of the primary sensory cortices and the motor strip. Terra incognita, at the time. “So how’s the comparison done? Bigger versus smaller?” Inhibition serves to subtract one input from another. If you were to listen in on a spinal cord neuron, you would discover that it was excited by a small patch of skin, but inhibited by an even larger patch of skin surrounding the excitatory patch. From the one neuron’s point of view, the world consists of two areas of skin that contradict one another. In the business, their combined skin area is known as the neuron’s “receptive field.” “That’s something like a city’s watershed, up in the mountains?” I nodded agreement. And for a neuron back in the spinal cord or brain, the watershed has two antagonistic regions, one sending hot water and the other cold water, mixing in the neuron to determine how hot it acts. So any one neuron to going to have a limited knowledge of what’s happening out on the skin. Collectively, a number of such neurons can form a committee that says “Watchband” from the temporal and spatial characteristics of their activity. Sensory neurons are usually excitatory at all their axon terminals — but some end on inhibitory neurons that, in turn, connect with the neuron doing the comparisons (making it both a second-order and a third-order neuron; the convention is to name it for the shortest possible path from the sensors). Each sensory axon from the skin has — at a minimum — tens of thousands of axon terminals. Since a neuron has a threshold for initiating impulses, it may be quite silent unless the excitatory input starts to considerably exceed the inhibitory input. If it is only inhibited because the skin stimulation avoids the excitatory patch, the neuron doesn’t pass on the information. “So the uniform pressure of a wristwatch band may be ignored by many neurons in the spinal cord, simply because inhibitory patches cancel out the excitatory ones?” Doesn’t cancel out in all of them. A dull pencil point, however, would keep lots of them active. And would likely result in a sensation being transmitted to higher brain centers, perhaps eventually reaching a level where it could be talked about. But, of course, a decision to act can be made by your spinal cord before the brain even knows what’s up. The neurons down there, on their own, are likely to command some muscles as a result of the sensory information — without waiting for the “higher centers” to comment. A quick withdrawal from a threatening stimulus is underway even before the brain knows about it, as when you step on a thumbtack. And the many modifications of routine body movements for walking or maintaining posture are made by the spinal cord without much need for involvement by higher centers. Similarly the second neuron in the pathway from the ear, located in the brain stem, may command protective reflexes against loud sounds without awaiting cortical intervention. But the reflex pathway always involves a round trip into the brain or spinal cord and then back out. “You mean that a muscle doesn’t contract because a local sensor directly commanded it? Why’s that? Seems it would be faster.” Faster, perhaps, but full of problems. Invertebrates sometimes use local circuits in the periphery, but many actions demand an integrated response of many parts of the body. Even if you step on a thumbtack, you may not want to withdraw that leg. “Why ever not?” Suppose that your other leg wasn’t on the ground at the time. Initiating leg flexion in that situation would cause you to collapse — indeed, toward the source of the pain. Giving autonomy to the local sensors and muscles, to make their own decisions, can be dangerous. And so most reflex paths are more roundabout, the decision-making abilities concentrated in the central nervous system (a collective term for spinal cord and brain). But yes, that does indeed slow down reaction times. Let’s take some high-end sensory abilities. Sound and sight are the most varied of our sensations. Language, invented sometime in the last few million years, involves some really fancy abilities for categorizing sounds. But fancy visual abilities have been important for much longer, and it shows. The visual pathways in the brain seem to have elaborated and streamlined in ways that have no counterpart in the hearing pathways. Our primate ancestors of 40 million years ago may not have needed to read, but they needed to look up at the top of a tree waving in the breeze, causing patches of bright sky to flash on and off, and nonetheless spot the little dark patches of fruit amid the dark leaves. Having identified the form of fruit, they needed a sense of color to judge whether the fruit was ripe enough to make it worth climbing up to the top of the tree. “I certainly climbed an apple tree a few times when I was a kid, only to discover that the apples at the top really weren’t ready to eat. Now I just wait until they’re surely ripe.” But you probably don’t have other primates raiding your apple trees. The monkey needs a fine sense of color, capable of discerning almost-ripe fruit. If the monkey waits until it’s unequivocally ripe, some other monkeys will have picked the tree clean in the meantime. Life in the trees also shaped up our abilities to operate vehicles. Swinging through the trees produces a visual stream rushing past on both sides of your head. And so does riding a bicycle, with many protruding branches to track and avoid. Some objects move faster than the average visual stream, and so are judged to be closer — or perhaps even moving themselves, such as other monkeys in the trees. We largely owe our excellent visual abilities for judging form, color, distance, and motion to the monkeys of long ago. Whenever we drive a car or read a book or admire a sunset, we are using neural machinery that was honed on the difficult task of finding ripe fruit amid clutter and confusion. Yet this neural processing is imperfect: we sometimes see things that are not there, sometimes fail to see things that are really there, and often distort things. Our visual system is not a video camera. 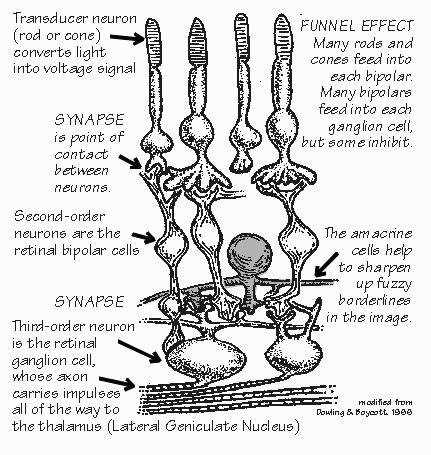 [FIGURE 52 Transducer neuron, retinal layers] “Of course, at first glimpse, our eye is rather like a camera.” Yes, in the sense that the lens produces an inverted image of the world on the back of the eye. Rather than the little silver grains of film that turn black when light strikes them, our retina has a great mosaic of 100 million photoreceptors that change their voltage, not unlike those in a very-high-definition television camera. But the analogies stop there. Our visual system is in the business of taking apart the visual image, not preserving it for some little person inside the head to view. |
|
UNLIKE MOST OTHER SENSES, the comparisons between neighbors in the visual system
start right out next to the sensor layer. “I thought you just said that comparisons are always made in the central nervous system.” Quite right. But the eye’s an odd exception because the retina is part of the central nervous system, just as the spinal cord is. The brain actually grows out into the eye during fetal development. The optic nerve is really only an honorary nerve — properly speaking, it is part of those internal tracts that make up the white matter of the brain. Each of the third-order neurons in the retina, whose axons make up the optic “nerve,” receives input from thousands of photoreceptors but never directly, only via intermediate cells called bipolar cells and amacrine cells. It’s like a funnel that collects raindrops from a wide area and concentrates them into a narrow stream. There has to be some funneling, as there are about a hundred photoreceptors for each axon going back to the brain. “Sounds like the visual system discovered what, in computer design, is called a preprocessor. They’re handy to reduce the amount of information that must be transmitted over long distances. So they do a preliminary analysis and only ship out the results.” The funnel isn’t quite the right analogy, actually. It isn’t a matter of a hundred photoreceptors connecting to each ganglion cell. Messages from thousands of photoreceptors are funneled into each ganglion cell, except some cancel out the actions of others. That’s because each photoreceptor contributes to hundreds of ganglion cells — there’s a spreading out as well as a funneling. There’s a hundred funnels beneath each photoreceptor. 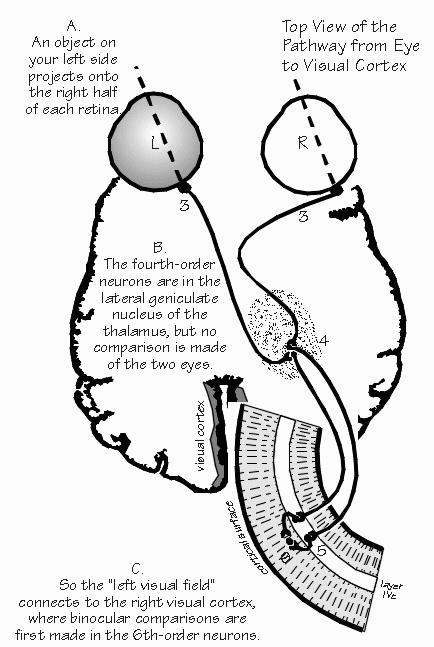 [FIGURE 53 Top view of the path from eye to visual cortex] |
|
FROM EYE TO BRAIN, the route goes through the thalamus, whose lateral geniculate nucleus
is an elaborately layered structure. Surely, everyone thought, something fancy happened in those
six layers, all of which send axons to the visual cortex. Three layers receive inputs from one eye
and three receive from the other. “Why?” It still isn’t known. But let me try asking a question that eye surgeons like to pose to medical students: What is the most important reason to save a damaged eye? “Wider field of view with two eyes?” But that surely isn’t the most important reason, as we can get around pretty well with one eye patched. There is a crescent-shaped region seen by one eye that is not seen by the other. We just have to turn our head or good eye a little to see it, if missing the one eye. “The range-finder effect, comparing the view from the two eyes to judge how far away something is located?” While we obviously improve our depth perception with the range-finder effect, that too can be mimicked by moving the head sideways momentarily, as when we shift our weight from one foot to the other. That gives us two views to compare. There are various other indicators of an object’s distance as well, such as how much surface detail you can see. No ability is really lost when we lose one eye. Here’s the answer to this Socratic question about saving a damaged eye: In case the remaining eye is damaged at some future time. Blindness is a very serious business. Any threat to the vision in one eye is far more serious than the immediate problem. And sometimes the threat doesn’t involve overt damage to the eye, just lack of sufficient experience. For example, I saw a boy named Ross in one of the teaching conferences. A nice six-year-old who is very cross-eyed. And now he’s almost blind in his right eye. The eye wasn’t damaged — it’s just that his brain ignores anything coming from his right eye and relies totally on the left eye instead. “But why?” That’s not known. Unfortunately it’s now too late to do much about this “amblyopia.” Cross-eyed babies who see doctors earlier in life get sent to see the eye surgeon, as doctors know that the child is very likely to become functionally blind in one eye or the other. Fixing the squint (usually in the first year) prevents the loss of vision in one eye or the other. Lack of regular well-baby checkups is not the only reason that Ross is now blind in one eye. His mother also thought of correcting crossed eyes in the same cosmetic terms as fixing a crooked nose. A perfectly understandable common-sense notion — but wrong. There are critical periods in the development of the brain where the right experiences are needed, or things get set up wrong — and then get set in concrete, as it were. 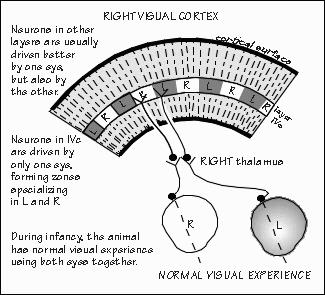 [FIGURE 54 Cortex’s ocular dominance columns in normal visual experience ] “And normal depth perception is one of them?” Well, maybe not depth judgments using the two eyes — maybe just the two eyes getting used to working in tandem, so you can fuse the two images. That one shouldn’t wait around with a cross-eyed baby, or bandage an eye for very long, were just empirical facts in medicine until the 1960s, along with the fact that nothing terrible happened if you patched an adult’s eye for equally long. Empirical just means, “Don’t know why, but that’s what happens.” There was something different about infants. Finally, research on kittens and infant monkeys advanced far enough to show us some reasons. 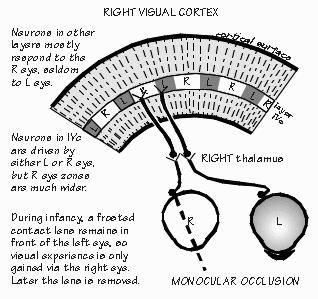 [FIGURE 55 Ocular dominance columns when one eye patched ] The two eyes seem to need experience working together at an early age. Now you’ve got to know at what level in the visual pathway the neurons begin comparing the slightly different view from the two eyes. It’s not until the sixth-order neuron. The fifth-order neurons — the ones in cortical layer IVc which receive the lateral geniculate inputs — are still “monocular,” just as were all the neurons in geniculate: if the neuron responded to the right eye, it wouldn’t respond to the left eye. But when recording from sixth-order neurons in the layers above and below the fourth layer, many neurons are found that respond to both eyes. Usually the receptive field for the left eye is similar to that for the right, but the response is usually stronger for one eye than the other. For some sixth-order neurons, the right eye is the “stronger” one, and for others the left eye has the most influence. Only a few are strongly biased toward one eye and ignore the other. But in infant monkeys wearing an opaque contact lens, this changes. Suppose the opaque contact lens is switched every day from one eye to the other, so that both eyes get visual experience during the first year of life — but never together. Recordings made from the brains of these animals are very different from normal ones. Now the neurons have strong preferences for the left eye or the right eye, with very few neurons paying attention to both eyes. And even if the contact lens is then left off for many months, so that the two eyes get a chance to work together, this bias tends not to reverse: few of these biased neurons drift into comparing the two views. “So the bad eye has been disconnected?” That’s probably too strong a way of expressing it, since drugs that block inhibitory synapses can temporarily reveal some underlying connections from both eyes. But for all practical purposes, the functioning of the visual system seems to have been permanently altered by this abnormal early experience. 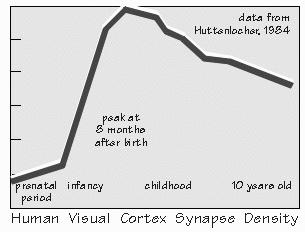 [FIGURE 56 Pruning of human visual cortex synapses during childhood] “But what causes a critical period?” Probably a lot of disconnections. Even in normal people, a progressive reduction in synaptic connections occurs with age, all over the cerebral cortex — it seems that things are connected up somewhat indiscriminately to start with, then refined. You had the most synapses per neuron that you’ll ever have back when you were eight months of age. After that, it’s all downhill — you lose a third to a half of cortical synapses during childhood. “That’s amazing. We’re losing connections while we’re storing additional vocabulary, accumulating more memories?” Yes — at least, on the average. In addition to detaching synapses, reduced connectivity also occurs by neuron death in some cortical regions. A monkey’s motor cortex loses a third of its neurons during infancy and the juvenile period. “They eliminate the clumsy ones?” That might be a good pruning principle. In any event, relatively few cortical neurons are lost after reaching adulthood. The current thinking about pruning principles is that there is elimination of the less active connections during this “sensitive period.” That you retain those synapses that you use a lot. 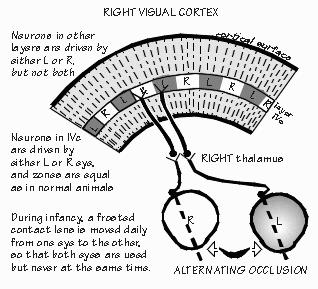 [FIGURE 57 Ocular dominance columns if patch alternates between eyes] A good example of this activity-dependent pruning principle is seen in layer IVc where the geniculate inputs terminate. Normally, there is a zone about 0.5 mm wide that is exclusively left eye, flanked by right eye regions that are also 0.5 mm wide. Suppose the opaque contact lens remains on the left eye, rather than being switched back and forth daily, so that only the animal’s right eye sees the world normally. What happens is that the right eye zones in visual cortex’s layer IVc expand at the expense of the zone for the unused left eye. And again, this is not readily reversed by leaving off the opaque contact lens once the critical period is over. “Sounds as if those exercised neurons had won a competition.” Yes, “use it or lose it” is still our best theory for why there are critical periods in development. And why kids such as Ross still can’t see in one eye, even when their squint is belatedly corrected. |
|
NEIL WAS VERY INTERESTED in the notion of synapses being pruned, presumably using
some pruning principle fancier than the one he used on his apple trees. “So are synapses being made and broken all the time?” he asked. Probably, although surely there is a core of fairly stable synapses. Remember all those experiments with rats living in enriched environments? They increased their number of synapses per cortical neuron by 80 percent. That shows you that the rate of creating synapses sometimes exceeds the rate of breaking them. The average decline with age, after eight months of age, shows you that the rate of breaking can predominate in the long run. But we don’t know if 10 percent are turning over every week, or every year. Until our research methods improve, we just know the results of the average differences in the rates of making and breaking. “It all sounds like economic theories, you know. And speaking of pruning, Joseph Schumpeter — an economist of about a half century ago — used to emphasize that `creative destruction’ was needed on an ongoing basis by society.” Certainly our memories are malleable enough to suggest that true events can be overwritten by false ones, like those eyewitnesses who came to believe they saw a yield sign rather than a stop sign. Year after year, we add more memories atop the old ones — and our brains aren’t like a file room that fills up and has to be cleaned out occasionally, but some different kind of memory system that carves a new pattern atop the old carvings. Occasionally the old patterns get pretty hard to see. “It’s like a sculptor getting carried away with details. If you carve away for too long, nothing’s left!” smiled Neil. But then he frowned. “Does that mean there’s a limit to how much our brains can store? If we live too long, do we run out of brain to carve patterns into?” Brains aren’t inanimate raw material like wood and stone. They renew themselves by making new synapses. And it’s the pattern of those synaptic strengths that seems to be the long-term memory trace. The question is whether you’ll continue to be able to retrieve some of the rarely used records, whether they’ll get overwritten so thoroughly that retrieval takes too long to be useful.
|
 Conversations with Neil's Brain:
Conversations with Neil's Brain: The Neural Nature of Thought and Language (Addison-Wesley, 1994), co-authored with my neurosurgeon colleague, George Ojemann. It's a tour of the human cerebral cortex, conducted from the operating room, and has been on the New Scientist bestseller list of science books. It is suitable for biology and cognitive neuroscience supplementary reading lists. ISBN 0-201-48337-8. | AVAILABILITY widespread (softcover, US$12).
|