|
Conversations with Neil’s Brain The Neural Nature of Thought & Language Copyright 1994 by William H. Calvin and George A. Ojemann. You may download this for personal reading but may not redistribute or archive without permission (exception: teachers should feel free to print out a chapter and photocopy it for students). William H. Calvin, Ph.D., is a neurophysiologist on the faculty of the Department of Psychiatry and Behavioral Sciences, University of Washington. George A. Ojemann, M.D., is a neurosurgeon and neurophysiologist on the faculty of the Department of Neurological Surgery, University of Washington. |
|
Taking Apart the Visual Image
NEIL HAD JUST GOTTEN the good news, that another patient had to be rescheduled. And so
Neil was on the O.R. schedule for the day after tomorrow. 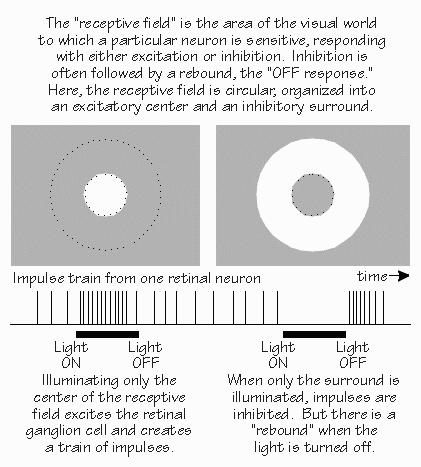 [FIGURE 61 Funneling to a retinal neuron for center-surround antagonism] In addition to antagonizing the excitatory response when a large light spot is turned on, the inhibitory areas of the receptive field tend to announce when a light leaves the inhibitory area. There’s a brief burst of impulses that says, in effect, “Light off.” “So a burst of impulses from that neuron could either signify the light turning on, or the light turning off?” Right. This third-order neuron makes, in effect, a temporal comparison in addition to its spatial comparison. Moving the light spot is capable of getting an otherwise silent neuron to perk up and take notice. While second-order neurons have some tendency to emphasize light-on and light-off, it is the third-order neurons that exaggerate conditions that change in time. Much of this was discovered in the cat retina by the neurophysiologist Stephen Kuffler in the 1950s, building on earlier work by H. Keffer Hartline in the frog retina. 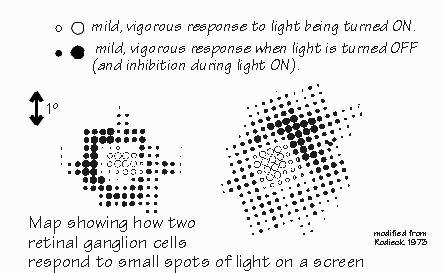 [FIGURE 62 Comparing two retinal ganglion cells] One reason that light-on and light-off is so important is that the visual image is rarely unmoving — it jitters around by the width of a few photoreceptors. So even if nothing is moving in your field of view, everything is in motion on the retina — even when you try to keep your eyes focused on a single point. The visual image is broken up into areas of near-uniform illumination and the boundaries between them. And it’s boundaries that activate so many neurons. When you stabilize the image on the retina with a fancy system that eliminates the jitter, parts of the image tend to fade out. “Really? You mean your view of someone’s nose might fade because it isn’t moving, but his hair might remain visible because it was blowing in the breeze?” Yes, a little movement is very important in keeping the brain interested in something. So the system produces some movement, even when the external world doesn’t have any. Thanks to this micronystagmus, the third-order neurons in a boundary zone between different shades of gray will “see” flickering light — from their point of view, and that’s what’s important here. Even somewhat fuzzy boundaries are detected by this system. Lines can look sharper than they actually are, because of this. So, even out in the retina, you see both of the major building blocks of visual perception, temporal contrast, and spatial contrast. |
|
THERE ARE FOURTH-ORDER NEURONS back in the brain stem, but the main path to the
cortex goes through the thalamus, its lateral geniculate nucleus. The fourth-order neurons there
behave rather like their third-order inputs from the retina — at least, when tested with
black-and-white stimuli rather than colors. Their receptive fields are the same center-surround
doughnut shapes as in the retinal neurons. “But I thought you expected to see big differences there?” Yes, it was quite a surprise back in the early 1960s when David Hubel and Torsten Wiesel discovered that the geniculate neurons were so similar to the retinal ones. We’d expected to see major transformations, but things turn out to be more subtle. The most noticeable difference is that diffuse white light is an even poorer stimulus for geniculate neurons than for the retinal neurons earlier in the path. While retinal neurons often exhibit some net excitation when uniformly illuminated, excitation and inhibition tend to completely cancel out in the geniculate cells. But that’s stimulating with white light or shades of gray. Try out colored lights and something new appears, not seen in the retina. The neuron may be excited by red light over a wide area, and inhibited by green light over that same wide area. 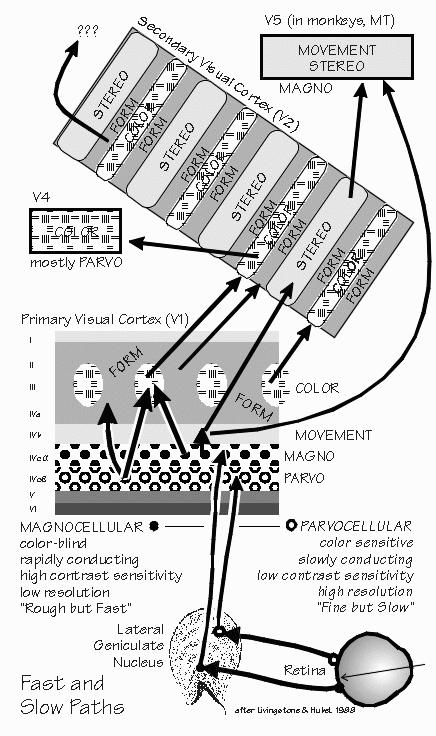 [FIGURE 63 Visual messages segregate into fast and slow paths] “No more center and surround?” In some geniculate neurons, the center-surround organization simply disappears. Besides this novelty, the geniculate turns out to have some differences between neurons in the bottom two layers and those in the top four layers. The top four layers have small cells (“parvocellular”) and the bottom two layers have noticeably larger cells (“magnocellular”). The magnocellular layers receive preferentially from the larger third-order neurons in the retina. “So they’re the fast track.” Not only that, but these magnocellular-layer neurons send their axons to cortical regions particularly concerned with small changes in spatial contrast and small movements. They quickly detect changes in the visual image and tend to exaggerate them. At least, they do this more than do the neurons of the four parvocellular layers, which receive preferentially from the smaller third-order neurons of retina and send their axons to regions of the cortex more concerned with color and the more obvious contrast differences. “Filling in such details can be done more slowly?” Perhaps. One of the fascinating things is that the parvocellular layers look normal in dyslexic children — but the magnocellular layers look disorganized, their cells shrunken as if neural development had taken a misstep. 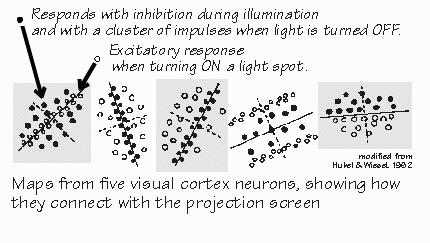 [FIGURE 64 Receptive field maps from visual cortex neurons] |
|
STILL FURTHER ALONG, the information from the eyes reaches the cerebral cortex. Here
things are really rearranged, with new principles becoming apparent. For example, the cortex
pays a lot more attention to elongated objects than it does to isolated spots of light. The cortex is no relay station in the manner that the geniculate was first thought to be. “Where did you get that term, `relay station’?” It comes from the old Pony Express, in the California land rush days, the places where the mailbags were switched from tired horses to fresh ones — but the messages weren’t analyzed or transformed. Geniculate, at first glimpse, is something like that. In the cortex, however, the messages are rearranged to make new patterns. The inputs to the cortex have those center-surround arrangements but are wired up to a cortical neuron in such a way as to create elongated receptive fields. The optimal stimulus for retinal and geniculate cells is a white spot on a dark background (or in other cells, a black spot on a light background) — always a round spot, although the optimal size varies among cells. In the cortical neurons, round spots might evoke a response, but the optimal stimuli are lines and elongated edges. 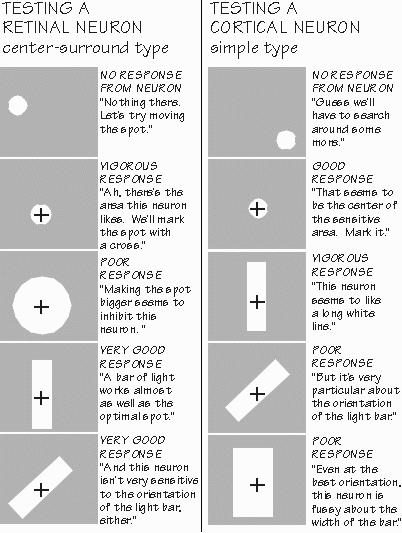 [FIGURE 65 Exploring receptive fields in retina and cortex ] “So you’ve got real line specialists. The frog’s eye may specialize in black dots like flies, but our visual cortex likes lines?” The emphasis on optimal stimuli can be overdone. If the stimulus is a line or an edge, of course, the inputs to the cortex aren’t firing at their optimum — which occurs with a spot of just the right size for them. So they’re responding only halfheartedly to the line. Yet a cortical neuron finds those inputs from suboptimally stimulated neurons optimal for itself. And so we have become cautious about characterizing the cortical cells as “optimal for straight lines” when they also respond pretty well to an eyebrow. Those fourth-order axons coming up to the cortex seem to be arranged so that the cortical neuron sums together the activity of many input cells whose receptive field centers were not all in the same place but rather strung out in a line. And so a white line on a darker background becomes the best way of maximizing the excitation while minimizing the inhibition. An edge (or a very wide line) may also be effective, as only half of the inhibitory flanks are thus stimulated. But diffuse white light that uniformly illuminates the receptive field is likely to cause inhibition to cancel whatever excitation gets through, as even the fourth-order neurons in the thalamus will fail to pass along the information in most cases (unless it’s a uniform color).  [FIGURE 66 Exploring the responsiveness of complex and end-stopped neurons] Some cortical neurons prefer horizontal white lines; others respond best to vertical ones. And there are specialists for the in-between angles as well. As you change the tilt of a line that optimally stimulates a cortical neuron, its response will diminish, disappearing within about 5 to 10 degrees away from the optimal angle. Of course, another group of cortical neurons become active at that point. “And I presume that some neurons prefer narrow black lines on white backgrounds.” Yes, as they get their inputs from those inverted Mexican hat receptive field neurons in retina and geniculate whose optimal stimulus is a black spot on a white background. Still other cortical neurons seem to like edges, such as a sky-sea boundary. The cortical neurons have a lot of variety, but it isn’t totally random. Neighboring cells tend to have the same preferences for tilt, until suddenly reaching a neighbor that prefers a quite different tilt. It appears that there is a column of about a hundred neurons in the cortex which are all organized similarly, at least for orientation preference. And that next door, there are adjacent columns interested in other orientations (often the “adjacent” angles, but sometimes jumping to an entirely different orientation). So we began to talk about “orientation columns” in the cortex as key features in the process of taking apart the visual image into important components. |
|
THEN CAME AN EVEN BIGGER SURPRISE: some cortical neurons would respond to the
optimally oriented line even when it was moved sideways. For many a lower-order neuron, that
maneuver would cause the neuron to be inhibited because the stimulus moves from the receptive
field center into a portion of the antagonistic periphery. There was a region perhaps 10 to 15
degrees wide where such a neuron still
responded to the line. But only when the line remained at its optimal orientation: try tilting the
line away from its favored orientation, and the neuron would stop responding. “The neuron is sensitive to orientation — but it’s independent of where the line is? That’s sure odd.” Hubel and Wiesel called these “complex cells” in distinction to the “simple cells” sensitive to orientation but also place. “So are the complex cells generalizing on the concept of line tilted at 45 degrees?” Psychologists have talked about generalization as a difference between lower and higher animals. Some species will learn to treat an erect and an upside-down triangle as “the same thing.” Yet other species will always treat them as different, refusing to “generalize on the concept of triangle.” Complex cells are generalizing, not about triangles but about one of the parts of the triangle, a tilted line. “This is all very interesting,” Neil said after finishing off his lemonade, “but it’s getting a little abstract. Do you see any patients with such patterning problems?” Every few years, someone reports a case of an epileptic whose seizures are triggered by patterned light, I told him. “Flickering lights, like those strobe lights they used to have in discos?” Sometimes, but in some patients a spatial pattern gives trouble too — as in looking at a screen-wire mesh covering the window or a herringbone pattern in a fabric. There was a case of two brothers who both had seizures triggered by spatial patterns. Their mother would sometimes find one of them frozen in place, staring at a screen door. Sure enough, when hooked up to the EEG machine to record their brain waves, fine repeating patterns would evoke the preseizure waveforms. But only if the lines in the pattern were tilted at a particular angle. It certainly seemed as if their brains got carried away when one type of receptive field was activated. |
|
SO THE VISUAL CORTEX has at least two new building blocks, oriented-line-in-a-place and
oriented-line-in-many-places. A third type soon appeared, the “end-stopped”
receptive fields where the neuron liked lines, but only if they didn’t exceed a certain
length at one end. Are there higher-order cells in the brain that specialize in triangles, no matter
what their size or tilt, no matter whether black-on-white or white-on-black, no matter whether the
center is filled or not? Not so far. Taking apart the visual image doesn’t stop in this first cortical visual area. It continues next door in the second, where specialized subregions emphasize form, color, and distance away. Indeed, there are nearly three dozen “secondary” visual areas so far — and that’s in monkeys. No one knows how many there are in humans, whose cerebral cortex is an order of magnitude more extensive than that of the monkeys. And, if that three-fold variation in size of the human primary visual area is any indication, there ought to be a lot of individual differences in these higher-order visual processing areas as well: inborn differences, plus all the variations due to different early experiences with the visual world.  [FIGURE 67 Generalized triangle category must handle many triangle varieties] “But there are no triangle detectors?”, Neil teased. “And here I thought that you were going to tell me about the neuron that specializes in my grandmother’s face. How are you ever going to get proper names for unique individuals if you can’t even get cells to represent common categories like triangles?” The neuron that is so specialized that it responds only to the sight of your grandmother’s face is, alas, a fictitious beast closely related in the cerebral mythology to the boss in the executive suite. We’re delighted with all the feature-extracting neurons of visual cortex — but we also doubt that such extreme overspecialization is always needed. If a committee of three generalists is good enough to distinguish between all the colors in the spectrum, maybe a committee of several dozen will suffice for all the faces you know.  [FIGURE 68 Trichromaticity from three cone types and the purple problem ] The “trichromaticity” committee principle was first discovered by Thomas Young, back in 1802. He was a British physician whose many interests included Egyptology. Then the principle was reformulated by the German physiologist-turned-physicist Hermann von Helmholtz in 1860, and fleshed out by the biophysicists who finally — about 1960 — got the techniques to see and measure the three cone types predicted by Young and Helmholtz. “Trichromaticity means that mixtures of red, green, and blue will mimic any color you want?” Neil asked. “The way it works on my color TV?” Right. One of the photoreceptors is more sensitive to the long-wavelength end of the spectrum — actually it peaks in the yellow-green, not in the red. Another has peak sensitivity in the middle wavelengths near green but has skirts covering the spectrum from blue to yellow. The third emphasizes the short-wavelength end, peaking in the violet but still somewhat sensitive to the yellows. With only one type, you don’t know whether its response is due to a bright yellow light, or a dim violet light. With three receptor types, you can sort out this ambiguity. With only two types, as in the common kind of color blindness, you will confuse colors in some situations. With three types of cones, you get a committee judgment that’s more reliable. “Is that where colors like purple come in? Purple isn’t in the rainbow.” Purple happens when you get strong votes from the long- and short-wavelength photoreceptors, at a time when the middle-wavelength photoreceptors aren’t saying very much. If the middle were to vote equally heavily, you’d see white instead of purple. If the long-wavelength signal were strongest, with the other two very weak, you’d call the light red. Note that when this happens, we call it red even though the only active photoreceptor actually peaked in the yellow-green. Red is a property of the committee votes in such a ratio, not of any one photoreceptor type. That’s why we’re so cautious about emphasizing the optimal stimulus shape for those visual pathway neurons. If color is any lesson, the peak sensitivity isn’t as important as the spread in sensitivity. And which combinations are activated together. That’s the lesson of color mixing, and we’re constantly rediscovering it. Taste works the same way. There aren’t any true specialists among the taste buds in the tongue, just four broadly tuned sensor types, all a little different. “Reminds me of what that gourmet cook is always saying on television, that good taste is a matter of getting the right combination of ingredients. I do wish that cafeteria cooks would learn about those interesting sweet and sour combinations, like strawberries and rhubarb,” Neil said. “I serve on a program committee,” he continued, “and I run into some of the same people when I go to a meeting of the fund-raising committee. So you think that, like me, a neuron is a member of a lot of different committees? And that the category is implemented by a committee and not a specialist neuron?” The Hebbian ensemble notion is that, when you recall a word like peanut, you are reactivating that particular committee of neurons. The committee may not look like a peanut, not any more than the bar code on a can of peanuts does, but it serves to represent the cerebral code for the peanut. A proper name, such as your grandmother’s, is likely to require a somewhat larger committee, so as to distinguish her from all the other women you know. “So it might take a dozen neurons to get the category, but another dozen to specify it further, narrowing it down to a particular individual?” That’s the idea, although it might be hundreds rather than dozens. The stroke patient who can’t recognize his wife’s face, even though he can tell one face from another, usually has a rather large area of damage on the underside of the temporal lobes. And such a patient has just as much trouble spotting a picture of his own car amid a series of photographs of similar cars. It’s as if he can’t narrow things down further, from the general to the particular. “But surely it’s important to know the basic elements,” Neil said. “You can figure out color mixing a lot better for knowing those three cone types. You can’t make much sense of purple without them. And as the TV cook says, you’ve got to know your basics in order to innovate, make the really interesting new combinations.” Yes, and the closer we get to knowing the shapes and colors and motions that the individual neurons like, the better we can understand something like recognizing faces, and why we make the mistakes that we do. “Mistakes like forgetting my seat belt, fifteen years ago?” Sorry, we don’t know the origins of major mistakes yet. But minor mistakes, such as using the wrong word — that we’re starting to understand. Tomorrow night, before the operation, we’ll give you a preview of the slide show you’ll see in the operating room. “I’ll probably make lots of mistakes on that test. George said he could almost guarantee it.” Yes, he can. |
 Conversations with Neil's Brain:
Conversations with Neil's Brain: The Neural Nature of Thought and Language (Addison-Wesley, 1994), co-authored with my neurosurgeon colleague, George Ojemann. It's a tour of the human cerebral cortex, conducted from the operating room, and has been on the New Scientist bestseller list of science books. It is suitable for biology and cognitive neuroscience supplementary reading lists. ISBN 0-201-48337-8. | AVAILABILITY widespread (softcover, US$12).
|