|
Conversations with Neil’s Brain The Neural Nature of Thought & Language Copyright 1994 by William H. Calvin and George A. Ojemann. You may download this for personal reading but may not redistribute or archive without permission (exception: teachers should feel free to print out a chapter and photocopy it for students). William H. Calvin, Ph.D., is a neurophysiologist on the faculty of the Department of Psychiatry and Behavioral Sciences, University of Washington. George A. Ojemann, M.D., is a neurosurgeon and neurophysiologist on the faculty of the Department of Neurological Surgery, University of Washington. |
|
The Personality of the Lowly Neuron
THE “GRAY” MATTER OF THE CEREBRAL CORTEX is just a thin sheet atop the
white matter. But within that reddish brown icing on the white cake, you can see the neurons
arranged into a half-dozen layers. In this case, it’s the icing that has the layers, not the
cake itself.
“That’s a simple flowchart,”
Neil said, looking up from the sketch on his napkin. He’d just come from his latest
clinic appointment, after taking what he hoped was the last of the neuropsychological tests.
“So, have you got the circuit diagram all worked out by now?”
All together, there are 148,000 neurons beneath a square millimeter of cortical surface, roughly a hundred million per square inch. They are often organized into little minicolumns of about 100 neurons apiece. Those in turn are sometimes organized into macrocolumns of perhaps 300 minicolumns with 30,000 neurons. “Sounds like a modular architecture to me,” Neil said, his engineering background resurfacing. “Almost like a housing development. So what do the elementary modules do?”
You’d think we could figure out a modular wiring diagram from all this information, but it’s surprisingly difficult. In the sensory strip, you’ll see a macrocolumn specializing in skin sensations, but the adjacent one might specialize in reports from the muscles and joints. In the visual cortex, a minicolumn will specialize in lines of a certain orientation, while adjacent minicolumns specialize in other angles. And the macrocolumns there are organized by eye — first you might see a set of minicolumns favoring the inputs from the left eye, then encounter another macrocolumn next door that favors what’s coming in from the right eye. But for most cortical areas, we don’t know much about the columnar organization. “Don’t different cortical areas look different under the microscope?”
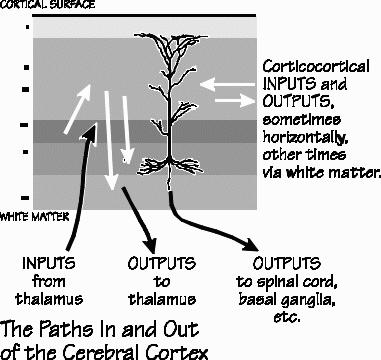
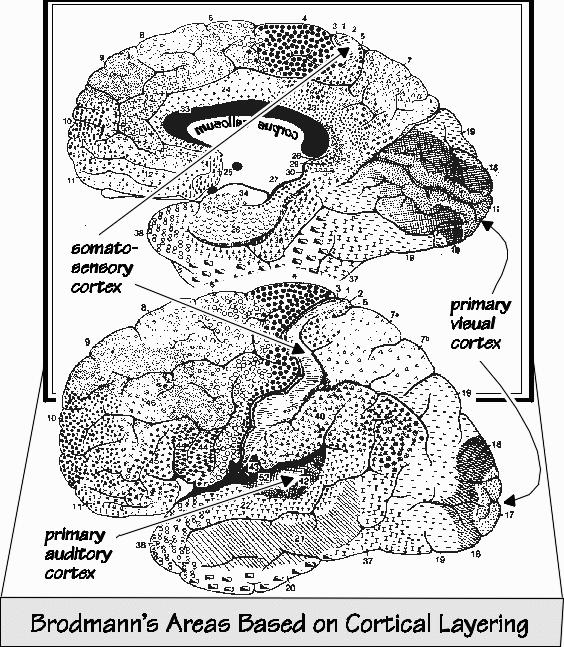
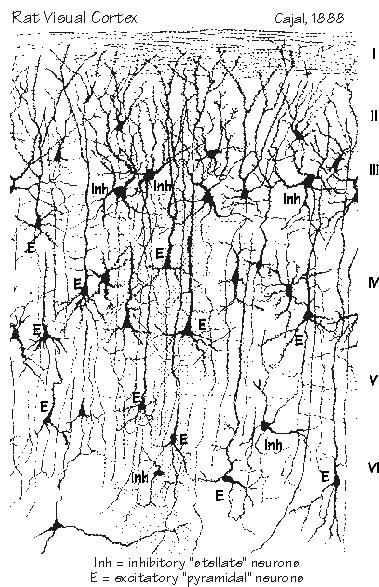
The half-dozen layers vary in their thicknesses. But the total cortical thickness stays about the same, all over the brain. Just imagine a city with a height limit of six stories for its buildings. But the ceiling height isn’t equal for all the floors — you might see a ceiling on a ground floor that is higher than the ceilings on the other floors. In one part of town, suppose buildings were constructed with the third floor extra tall for some reason, but with the fourth floor having a ceiling so low that you hit your head. Early twentieth-century neuroanatomists defined 52 cortical areas in that way, using neuron size and the relative proportion of layer thickness, and gave them identifying numbers. Area 17, for example, is otherwise known as the primary visual area. In the motor cortex, also known as Area 4, there isn’t much of a fourth layer because it doesn’t receive much information coming up from the thalamus. Yet back in the primary visual cortex, the fourth layer is the most impressive of all. That’s because it has so many inputs from the eye (via a relay in thalamus) that a lot of extra neurons are found in the fourth layer, bringing the total up to 357,000 neurons under a square millimeter. Because the extra neurons are densely packed, and the axons from the thalamus are so segregated, you can see a horizontal stripe in the fourth layer, even without a microscope. At the edge of Area 17, where it abuts Area 18, the stripe abruptly stops — that’s why the primary visual cortex was originally known as “striate cortex.” Unfortunately the language cortex isn’t as obvious in structure.
|


|
BACK WHEN SIGMUND FREUD was peering through a microscope, the gray matter looked
like a great spider web of crossing axons. Everything seemed fused together. At some of the
intersections, instead of a trapped fly, there was a black bulge, the cell. No
“arrows” anywhere, suggesting how information flowed. Freud probably found
this rather frustrating, and by the time visualization techniques were improved, he had moved on
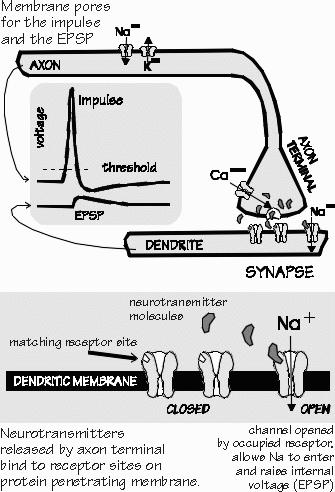
to view the brain from a very different perspective — psychoanalysis. With Camillo Golgi’s method of staining a few neurons at a time with silver, Fridtjof Nansen and Santiago Ramón y Cajal independently deduced that the axon actually came to a dead end just outside another cell — that there was really a gap, rather like the no-mans-land between the two independently maintained border fences of neighboring countries such as Israel and Syria. That was back in 1888. About 1900, the neurophysiologist Charles Sherrington gave this point of near-contact a name — the synapse. But the synapse wasn’t seen in more detail until 1953, following the advent of the electron microscope that showed those parallel border fences. The synapse is the border-crossing point, where the output of one neuron’s axon becomes the input to another neuron’s dendrite or cell body. What crosses this border is information, in the form of chemicals. It’s analogous to opening a bottle of perfume at one border fence and letting the molecules waft across the no-mans-land (the synaptic cleft) to the other border fence (the cell membrane). When reaching the other side, the perfume is “sniffed” by special receptor molecules embedded in the cell membrane. These “perfume molecules” are the neurotransmitters. There are dozens of different types, such as norepi and dopamine. A common one in the cerebral cortex is glutamate. Some neurotransmitter types are released at a considerable distance from the target neurons and diffuse around a neighborhood, rather like cooking aromas. They change the excitability of target neurons in a general way. This is called “volume transmission.” “Sounds like hormones.” Hormones are the long-distance version of this principle. The hormone is released at a distant site, such as the kidney, and carried by the bloodstream to its target cells, such as a muscle. The other extreme of the principle is the traditional synapse seen in the electron microscope, where the neurotransmitter is released so close to the target cell that it can’t miss — it has little chance to diffuse away and affect any other neurons. But there’s this intermediate version, where the molecules reach a whole neighborhood of target neurons. “A tonic of sorts? They get in those neighborhood neurons and turn up their volume control?” They can certainly change neuron properties, so that bursts of impulses are produced more easily. Of course, the neurotransmitter usually doesn’t enter the “downstream” neuron. It only tickles its surface receptors.
“It’s not absorbed? So what happens to the neurotransmitter?” It gets recycled, much of it back into the presynaptic side of the border fence. Many drugs interfere with this recycling of transmitters back into the cells that release them. We call the interference “blocking,” but it is closer to slowing down, less like a strike of the garbage collectors than a work-to-rule slowdown. For example, cocaine blocks recycling at synapses using dopamine as their neurotransmitter, among other actions. Common antidepressant drugs often block recycling at synapses using serotonin. It has long been known that many insecticides block recycling at acetylcholine synapses. “What’s the purpose of slowing down recycling?” So the neurotransmitter can hang around longer. This fools the downstream cell. Blocking recycling usually makes the postsynaptic cell think extra neurotransmitter was released upstream — something that usually happens only when a more imperative message is being sent from the presynaptic terminal. Overdoing it, of course (as in the case of insecticides), can make the synapse fail to communicate the arrival of subsequent signals because, like a sticky doorbell button, the postsynaptic receptors never reset. The flow of information is one way, most of the time. It goes from the upstream “presynaptic” neuron to the downstream “postsynaptic” one: only one side has the perfume to release, and only the other side has the receptor molecules. And that defines one end of the axon as the output, the other end as the place where the inputs last have a chance to interact. While neurons don’t come with arrows attached, they are effectively one-way streets for this fast flow of messages. Back in Freud’s day, no one knew this — the neural network just seemed like one big web with no input-output organization, no uphill or downhill.
“So let me see if I’ve got this straight. Something releases a packet of neurotransmitter molecules into that no-man’s-land between cells. Some are recycled, and a few tickle the receptors on the postsynaptic side — and then they’re recycled too.” Neil thought for a moment. “Sounds as if neurons compute on some sweet-or-sour principle using cooperative or conflicting perfume?” Only very indirectly. Some of the minutes-to-days reactions of neurons are indeed chemical, but on the timescale of reaction times and speaking sentences, computation is largely electrical. Sweet is a positive electrical change, and sour is a negative one. The perfumelike process at the synapse is just a middleman. But it’s an adjustable middleman, thought to be the basis for learning and memory. The only way you can understand that adjustability is to look at the electrical aspects of the neuron that control computation and synaptic strength — how much electrical effect a synapse produces. And the aspects that move the message from one end of the neuron’s axon to the other. |

|
ELECTRICITY CARRIES THE MESSAGE, except at the synapse itself. The event that usually releases the neurotransmitter, like a puff from a perfume atomizer, is the arrival of a puff of electrical charge. Though only a tenth of a volt from bottom to top, the “impulse” is the biggest voltage change seen in the brain. It moves like a wave from the input end of the axon to the output end — except in insulated axons, where it seems to hop between the gaps in the myelin insulation. “So this electrical signal doesn’t move almost instantly, as in wires?” No, this is more like burning fuses. The speeds vary among axons, ranging from the equivalent of the slowest freeway traffic jams for the smallest uninsulated axons, up to those of the fastest trains for the largest insulated axons. The impulse lasts only for about 1/1000 of a second at any one spot (a millisecond is also about as long as a fast camera shutter remains open), but that’s long enough to trigger the release of some prepackaged neurotransmitter, emptying it out into the synaptic cleft. And what happens on the other side of the synapse is also electrical in many cases. The neurotransmitter sticks to the receptor molecule, and together they are able to open a pore in the cell membrane. An electrical current then flows into (or out of) the postsynaptic neuron for a short time, producing an event known as a postsynaptic potential. All this can repeat, many times each second, depending on the impulse initiation rate. “Whoa! What started the impulse?” Postsynaptic potentials in the dendrites spread to push the voltage past threshold at the beginning of the axon, near the cell body. It’s like a trigger on a gun, an unstable equilibrium. “I’ve never understood where the electricity comes from.” One use of the food you eat is to charge up your batteries. Each cell in your body has such batteries, created when their cell membrane throws certain kinds of salts out of the cell and captures other kinds to hoard inside the cell. All this takes energy. Typically, sodium ions are excluded and potassium ions are hoarded. That’s the battery part. All of the interesting events, like impulses, involve making currents flow, using those batteries. When the flow of sodium into the neuron is smaller than the outward potassium flows, the voltage simply heads back to the resting potential. Just imagine a mixing faucet with a mind of its own: whenever something turns up the hot water, it turns up the cold water even more, until the water stream returns to some standard lukewarm temperature. But suppose your faucet is also designed such that, when the temperature rises beyond some trigger value, the hot water comes on much harder than the cold, and you get a blast of hot water before any compensation occurs to cool things off. The threshold is an unstable equilibrium: slight increases in the voltage above the threshold, and it’s off to the races. “I know some hotels with showers like that.” Their faucets aren’t that diabolical, only sticky. That blast of sodium ions entering the neuron is the impulse starting. Should the inward sodium flow exceed the potassium reactions, then voltage rises — and even more sodium comes in, and the voltage rises even further. It’s a vicious cycle. Eventually it stops at the peak of the impulse, where the potassium flow has finally increased enough so that the two flows cancel each other out. “But what stops the impulse? Why doesn’t the voltage hang up there?” That’s because the sodium pores tend to slowly shut themselves off at the higher voltages. The potassium pores don’t do this, so they eventually win. So these pores (we usually call them channels) open or close, depending on the local voltage. They’re quite different from the neurotransmitter-sensitive pores that produce the synaptic potentials and the initial rise of the impulse. The voltage-sensitive pores then take over to produce the rest of the impulse. “What makes the impulse travel down the axon so slowly?” This is something like the Pony Express passing on the message to a new horse and rider, the impulse has to be recreated next door, and next door to that. “Ah! The domino theory is alive and well — in neurophysiology!” Very much so. The fatty insulation wrapped around the axon has gaps, called nodes of Ranvier, about a millimeter apart — and the impulse seems to jump from one gap to the next. That takes time, more in some axons than in others — just as it takes time for each domino to fall. And the long axons may have thousands of such dominos. The impulse is repeated all the way down to where the axon branches. Then impulses travel down both branches, and so on, until shifting the presynaptic voltage at a thousand different synapses. That’s why the mass-mailing analogy is appropriate: the same message — say, a pair of impulses — is sent to all the recipients. “But what’s to prevent an impulse from traveling backward along the axon? Just as there’s nothing to keep me from driving the wrong way up a one-way street?” Not much. Of course, an impulse has to get started somehow — and normally the only place they start is at the beginning of the axon, near the input synapses for that neuron. But impulses can start anywhere, given enough impetus. You can start them from the middle of the axons that wrap around the back of the elbow, for instance. Sometimes this even happens with little provocation, as in the people with slipped disks that traumatize the nerves to their leg. “About your lawn-sprinkler metaphor — do those impulses starting at the end of the axon have something to do with the spread of the medium-sized seizures to cause a big seizure?” That’s one idea, and it’s also a possible cause of muscle cramps. Since the anatomy is simpler for the motor neuron and the muscle, it’s better to start there. Each time a motor neuron back in the spinal cord fires an impulse, the muscle fibers also repeat the impulse and thereby twitch (the synapse between axon and muscle is so strong that it always creates an impulse in the muscle cell). For some of the hip-and-leg muscles, the motor neuron’s axon branches enough to connect with several hundred muscle cells. Thus, the impulse in the motor neuron’s axon sets off the twitches in many muscle cells. A series of twitches builds up muscle tension into a sustained contraction. Now suppose something set off an impulse in one of the many axon terminals — which ordinarily would never happen. Just changing the environment around the terminal branch of the axon could do it; maybe it gets a little starved for oxygen because blood flow is insufficient. So off the impulse goes, the wrong way up that one-way street, heading back up to the spinal cord. “That must confuse the poor spinal cord.” Probably not very much. The problem is that this wrong-way impulse sets off a muscle contraction in each of those several hundred muscle fibers. Each time the impulse travels past a branch, going off to another muscle fiber, an impulse starts down that branch. And sets off a twitch. So some fraction of the whole muscle gets a little twitch. A single twitch occasionally may be irritating, but it’s not a serious problem. But should that abnormal site of impulse generation set off repeated impulses, then it will be just as if the motor neuron back in the spinal cord sent a whole series of impulses down the axon in the usual way. A sizeable contraction now develops. “And so I get a cramp that just won’t stop,” Neil said. “It’s totally out of control. At least, out of my control.” Right. Except for massaging the muscle or stretching it out by contracting other muscles, there’s nothing you can do about it. The brain just doesn’t have any pathways to where the trouble is located that can possibly affect this erroneous barrage. Willpower doesn’t work anymore, because the impulses aren’t starting inside the brain or spinal cord. “This is what happens when one of my medium-sized seizures suddenly generalizes into a full-blown grand mal?” That’s the idea. The neurons in the brain have axons with a lot of branches going different places. And so a wrong-way impulse generated by massive environmental changes in one place — and seizures dump a lot of potassium out of neurons — can travel backward and set off impulses going forward down each branch that it passes on its backward journey. If there’s a real barrage of wrong-way impulses similar to those preceding the muscle cramp, then you can get big effects that are widespread in the brain — both sides, arms and legs. And virtually simultaneously. “That happens only for the axons of the selective attention system?” No, it probably happens for many of the axons entering and leaving an area where there is a medium-sized seizure. An average cortical neuron has an axon with perhaps 10,000 side branches. But a neuron in the selective attention system or the brain-stem arousal system has a lot more side branches than average — maybe a million — and so it amplifies the mischief more effectively. “You said that the anticonvulsant drugs helped to stop this spread?” Either that, or the drugs make the distant sites more resistant to starting their own seizure when the backward barrage arrives — say, by increasing the stockpiles of inhibitory neurotransmitters so that they can’t be exhausted quite as easily when such a barrage invades. It takes a lot of wrong-way impulses to start a seizure in normal cortex. Just as with starting seizures with electrical stimulation, a lot of synchronized input — that keeps repeating for a while — is capable of driving any cortical area into a seizure. |
|
INTERFERING WITH NEURAL TRANSMISSION is the more classic way to influence the
flow of information within the brain. Local anesthetics may block impulse replication in mid-axon. Muscle paralyzing agents tend to interfere with the synapse between nerve and muscle.
Some agents like curare plug up the receptors that guard the postsynaptic pores, so that
acetylcholine can’t bind. Other agents prevent recovery after an impulse arises, and so
interfere with replicating the succeeding impulses. Furthermore, the ions that enter the postsynaptic neuron through the opened pore may do more than just change the internal voltage. The calcium that enters through some synapses, for example, may change a variety of regulatory processes inside the neuron. “It has a dual role in life?” It sends a second message, in addition to the voltage change it produces across the cell membrane. Some synaptic actions don’t involve membrane pores or voltage changes at all. Instead, the neurotransmitter binds to a receptor molecule on the outside of the cell membrane. This G-protein then manages to cleave a high-energy chemical bond inside the neuron, something of a trip wire that sets in motion various chemical reactions — some even send messages to the cell nucleus, affecting the manufacture of proteins. This is a much slower process, sometimes producing voltage changes but sometimes merely modulating others, in the manner of pressing the pedals on a piano to modulate the sound produced by the keys. The calcium-blocker cardiovascular drugs attempt to alter these internal effects. While some of the actions are prompt, it make take weeks for the antipsychotic and antidepressant drugs to do their job. “And, I presume, some anticonvulsants take a long time to act.” Yes. But second messengers aren’t the only slowpokes on the scene. Many synaptic enhancements don’t involve the postsynaptic neuron, but rather act to increase the neurotransmitter available to stimulate it. “Such as those things that block recycling?” Exactly. Many antidepressants just slow down the reuptake of the neurotransmitter into the presynaptic terminal. |
|
EXCITATION AND INHIBITION have a range of connotations, even Freudian ones, but to
neurophysiologists they mean something close to addition and subtraction, or deposits and
withdrawals. The neuron doesn’t subtract a molecule of inhibitory neurotransmitter
from two molecules of excitatory neurotransmitter, however. The neuron avoids the
“adding apples and oranges” problem by adding the positive and negative
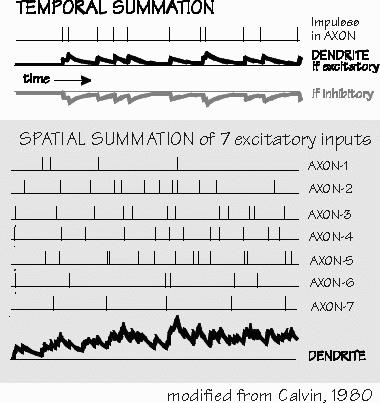
electrical currents that the neurotransmitters produce. “Ah! Electricity is the currency of computation in the neuron?” Instead of dollars, this currency is measured in nanoamperes of synaptic current. In the mixing faucet analogy, this would be the temperature of the water mixture emerging: the excitatory synapses “turn up the hot water,” and the inhibitory synapses turn up the cold. In general, all of the thousands of outputs from a neuron tend to have the same postsynaptic action, and so we can often speak of inhibitory neurons and excitatory neurons.
“Do they look different?” Often they do, at least in the cerebral cortex. There you can easily spot most of the excitatory neurons because they have a big “taproot” growing toward the surface called the apical dendrite. It arises vertically from the cell body (which is sometimes triangular, leading to the name “pyramidal neuron” for the whole class of taproot neurons). And, strangely, the apical dendrites of a dozen neurons may bundle together. The bottom of the cell body has some basal dendrites surrounding it like a ruff. The axon emerges from the cell body and dives into the white matter, en route to delivering a message far away. But the axon also has a series of side branches that end nearby. They are the most prolific source of cortical synapses in most areas of the cortex. Pyramidal neuron axon terminals are likely to release the excitatory neurotransmitters such as glutamate or aspartate. And the neurons downstream from them have receptors that can open up pores to sodium ions and so produce a voltage change known as the excitatory postsynaptic potential. But you can also raise the temperature by turning down the cold water in the existing mixture, and that’s how serotonin synapses from brain stem axons work in the cortex, by reducing the resting potassium current. “So they’re also excitatory. What about the inhibitory neurons?”
If you see a cortical neuron that doesn’t have the taproot design, it’s usually an inhibitory cell — and it usually releases gamma aminobutyric acid (GABA) from its axon terminals as a neurotransmitter. But what makes it inhibitory is the postsynaptic action of its neurotransmitter. “The cold water treatment, I presume.” What inhibitory synapses do is to oppose excitatory synapses elsewhere in the dendritic tree and cell body. The way this usually works, there’s a pore that doesn’t let sodium ions through, just mostly potassium or chloride ions. And so it produces an inhibitory postsynaptic potential that subtracts from the excitatory ones. About 40 percent of a neuron’s potential inputs, on average, are inhibitory. So it’s all a big balancing act. Excitatory synapses tend to raise the voltage above the impulse threshold and “fire” an impulse down the axon. But, individually, they are seldom strong enough to do this; it’s likely to be only a small percentage of the voltage change needed to kick off an impulse. “Except at the muscle synapse. Sounds like the brain is more subtle. And varied?” [RealAudio sound file (about 50k)]
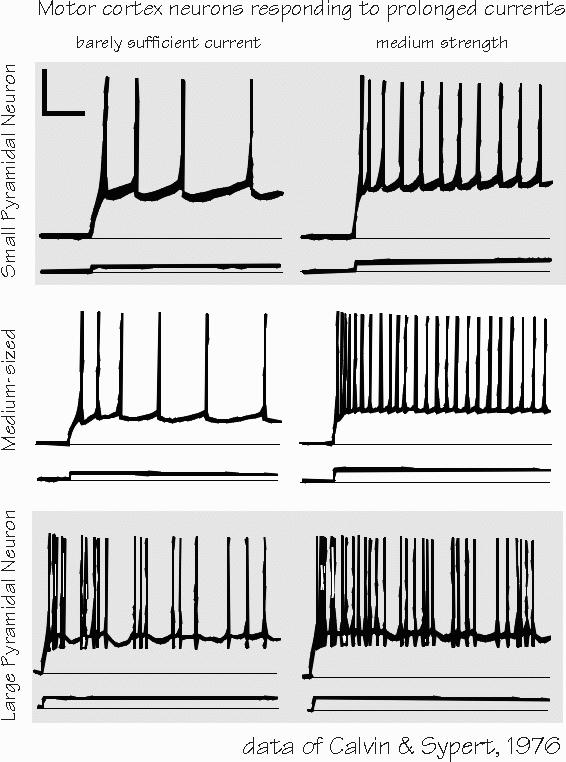
It’s almost as if different neurons had different personalities. Some are specialists in rapid change — they ignore maintained conditions. Others do the opposite. This turns out to be of major importance in dyslexia. You often hear it as an easy-to-remember dichotomy — “fast” versus “slow” cells, jittery versus steady, movement-sensitive versus static, magnocellulars versus parvocellulars. But there are lots of in-between types too, simply because there are more than a dozen types of pores into neurons, and the mix of these types (which is under genetic control and well as being affected by drugs and experiences) determines the neuron’s personality, just as different blends of whiskey determine its taste. Cortical neurons are often silent — George often sees human temporal lobe neurons that respond at rates slower than once per second. When neurons become active, they need more oxygen and glucose. They have ways of asking for more blood supply and, since we have nice ways of measuring that increased blood flow with PET scans and functional MRI techniques, we can produce colorful images of the brain at work. |
|
THE AMOUNT OF NEUROTRANSMITTER RELEASED may vary among the thousands of
output synapses. Another way in which the synaptic “strength” is varied is to
change the number of postsynaptic receptors to the neurotransmitter. Or you can change the
recycle times for the neurotransmitter. For these and other reasons, the synaptic potentials produced in the many postsynaptic neurons are not all of the same strength. While the synapse is only an inefficient chemical middleman in what are otherwise efficient electrical processes, it is a malleable middleman. Without this adjustability, the neural circuitry would be as rigid as that in our consumer electronics. The ability to adjust these “synaptic strengths” is what learning and memory are all about. In the short run, synapses change strength with use. Sometimes synaptic potentials decline in size and we speak of synaptic depression. “That’s depression?” Not the psychiatric use of the term, but an unrelated use of the same word by physiologists. And when one impulse follows closely upon another, there may be more neurotransmitter released by the second impulse than by the first — that’s called synaptic facilitation. Following a burst of impulses, any impulse in the next minute or so may release more-than-standard amounts of neurotransmitter. That’s known as post-tetanic potentiation, or when even longer times are involved, long-term potentiation — enough of a mouthful that it’s usually abbreviated LTP. “So synapses are what’s changeable. Now we can talk about how learning and memory works?” Such impatience. Well, just as a preview — temporary changes in synaptic strength such as synaptic depression and facilitation probably underlie our fading short-term memories. And it is the short-term memories that may provide the basis for the construction of permanent memories. If you lose something from short-term memory, it never gets kept in a more permanent form. “Why’s that?” Probably because some short-term changes provide the scaffolding for making permanent changes, casting things in concrete. LTP is an interesting candidate — it’s seen in only certain malleable parts of the nervous system. Among them are the upper layers of cerebral cortex that manage all that interoffice stuff. And LTP is one thing that makes synapses grow bigger and stronger, changing the brain structure in the long run. It’s how your thoughts rewire your brain, so it’s a different brain tomorrow than it is today. INSTRUCTORS: You may create hypertext links to glossary items in THE CEREBRAL CODE if teaching from Chapters 6-8, e.g., <a href=http://www.williamcalvin.com/bk9gloss.html#postsynaptic>Postsynaptic</a> |
 Conversations with Neil's Brain:
Conversations with Neil's Brain: The Neural Nature of Thought and Language (Addison-Wesley, 1994), co-authored with my neurosurgeon colleague, George Ojemann. It's a tour of the human cerebral cortex, conducted from the operating room, and has been on the New Scientist bestseller list of science books. It is suitable for biology and cognitive neuroscience supplementary reading lists. ISBN 0-201-48337-8. | AVAILABILITY widespread (softcover, US$12).
|
revised November 1996